Guomin Chen1 and Junru Wu1,2*
1Institute of Ultrasound Engineering in Medicine College of Biomedical Engineering, Chongqing Medical University, Chongqing, P.R. of China, 400016
2Department of Physics, University of Vermont, VT 05405, USA
*Address for Correspondence: Junru Wu, University of Vermont, Burlington, VT 05405, USA, Tel: +802-656-8357; E-mail: jwu@uvm.edu
Dates: Submitted: 23 September 2017; Approved: 28 November 2017; Published: 30 November 2017
Citation this article: Chen G, Wu J. The New Trend of Human Virology. Int J Bioanal Biomed. 2017;1(1): 001-009.
Copyright: © 2017 Chen G, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
According to the traditional virology, the invasion of virus to the human beings is of exogenous nature, i.e., the source of invader is external. Therefore, we always search for the other species for possible source of virus when a viral epidemic happens to human beings. For example, when the SARS and Ebola epidemic occurred, we naturally looked for the possible original virus sources from civets, and apes etc. [1-4]. Up to our knowledge, no successful search has been clinically confirmed. Vaccines have played effective roles to prevent viral diseases, such as smallpox, measles, and polio etc. [5,6] , they are found to be ineffective in the battle against other viral diseases, such as HIV infection and HBV and Flu epidemic [7-9]. This also prompted us to explore the possible significant differences of effects of the mechanisms of vaccines with children and adults. The following questions need to be answered: (1) Are there any mechanisms or drugs that can eliminate viruses in the nucleic acids form within cells? (2) Has the immune response really eliminated the persistence of intracellular viruses [10]? It is also known that the tumorigenic mechanism is related to the persistence of virus [11]. In our opinion, it is premature to conclude that the HPV vaccine used to prevent viral diseases is an anticancer vaccine [12]. The above-mentioned issues suggest that the relationship between viruses and their hosts may need to be further studied and clarified. The primary purpose of this article is to argue that virus inherently comes from the origin of life. To determine “what position of viruses should be placed on the evolutionary branches of the life tree” is important for understanding the origin and its development of virus. We emphasize in this article that the origin of virus is of endogenous nature, once it find its host, it develops with the host along with the cell evolution. As a large group of basic species, the virus is always associated with its long-term host and not randomly distributed or diffused. The classification of human virology should be based on the interdependence between viruses and human beings. We believe that the above viewpoints will significantly change vaccine applications as well as the concepts of human viral micro-ecology.
Origin of the virus
The endogenous nature of virus and the origin of cells
It is a challenging and controversial task to define “what is the life”; different professionals have different definitions. Generally speaking, the life is a material system with capacity of self-sustaining and self-reproduction and evolution [13]. In the process of evolution of life, the spontaneous happening of membrane vesicles with bilayer hydrophilic lipid molecule in Primitive Ocean has created a closed internal environment for the development of biological macromolecules [14,15]. RNA is the only molecule in the chemical world which can naturally produce the functional behavior, the natural chemical reaction can evolve the highly effective enzymatic response by selecting the proper functional ribozyme from the random RNA molecules [16,17]. The base pairing is a natural feature of nucleic acid molecules, once the appearance of RNA replication enzyme in ribozymes catalyzes the replication of RNA, it indicates that the gene is originated from the abiotic chemistry, resulting a chaotic world of genes [18,19]. Then the emergence of RNA’s reverse transcriptase, an analogues of RNA replication enzyme, makes these isolated genes to be linked from RNA sequence into DNA sequence, these genes have been preserved in a structure of genome with more stability and more complex functions, it means that the gene world have been turned into a stable and orderly genome world [20]. The electromagnetic energy of light can split water molecules into oxygen and hydrogen with the aid of pigment molecules [21], the hydrogen ions can be converted into hydrocarbons by combining carbon containing compounds [22]. When a primitive photosynthetic center consists of pigment molecules is inserted into the double lipid layers of vesicles, which prompts the accumulation of hydrocarbons and hydrocarbon energy in these vesicles. It is a macroscopic phenomenon taking place in the pristine ocean under abundant sunlight surrounded by an atmosphere with high concentrations of CO2. In the RNA world, numerous functional ribozymes are selected by various natural chemical reactions, those random chemical syntheses then turn to be the efficient ribozyme catalysis. After the chloroplast genes and genome are induced by the photosynthetic function, the original photosynthetic center from casually composition becomes biosynthesis. The vesical with interior particles of bio-photosynthetic center behaves as the most primitive life form in the process of life evolution [23]. Meanwhile the appearance of RNA encoding protein induced by RNA genome is a magnificent chapter of life evolution [24], it has completed the transformation of the life information process from nucleic acid sequence to amino-sequence, further to structural protein. All genomes in the vesicles of primitive period are equal and independent, and once one genome links with genes both in quantity and functions which is sufficient to control and represent the characteristics of the whole vesicle; the cell genome starts to appear and the life then begins on the cell stage [25]. The concept of “Species” is created with different cell characteristics of different cell genome. Once a cell genome controls vesicle performance as a cell, the accompanying genomes at vesicle stage fall back to subordinate or parasitic positions. In the evolutionary process, those isolated genomes which conflict with cell genome are gradually excluded and eliminated, and others are kept in a symbiotic or parasitic state, since their functions do not conflict to cell genome or cells, such as genomes of chloroplast, mitochondrion and ribosome [26-28]. The original virus comes from these genomes in vesicles as the plasmid is related to bacteria [29]. Since the symbiotic genomes including virus develop within a limited space of cells under the pressures of cell genome, they can share enzymes in the cell’s biochemical system. These genomes have lost their developing trend by gene linkage to the large-scale as cells’, but they still can use the “copy” capacity. When they do, the copy function very often with “errors”, such as losing the unnecessary nucleic acid sequence for miniaturization, [30] or/and overlapping use of nucleic acid sequences by various genes [31] for survival. In a way, the refinement of sequence composition of viral genomes and the large gap both in size and functions between virus and cells are really reflecting the magnitude of viral evolution (Figure 1). A RNA genome has its own structure with more complex functions, for example, ribosome genome [32], which is extremely “ancient”, has still preserved for its “brilliant” and irreplaceable functions in evolution. Limited by its configuration and stability, the RNA genome has limited number of RNA chains. The ribosomal genome consists of 3-5 RNA chains. The Influenza virus genome 8 RNA chains [33], and the reovirus genome 12-14 RNA chains [34]. By the way, RNA virus has no any relationship with DNA virus with their respective characteristics of formation, structure, replication and evolution. Each species of cell has its own independent free genomes with their own origins, these genomes are shuttling between cells in the manner of cell division, fusion, secretion, phagocytosis, and budding. Thus, the structure and behavior of these genomes already possess the viral characteristics. They were precursor of the original virus for composing the endogenous virus group of corresponding cells. Following the higher multicellular species evolution of the life, symbiotic virus group would further differentiate according to the compatibility selection rule and re-distribute by cell differentiation performance, the suitable ones by adaptation gradually evolves into endogenous viruses in a corresponding tissue cells. Since the survival rule is strict cell-dependent, in the multicellular species, the cells with different characters can be accompanied by their own endogenous virus groups. The development of virus in multicellular species requires not only the shuttle diffusion between adjacent conspecific cells, but also the infective channels between individual species for crossing the skin, the lumen of the cavity, the circulating fluid, etc. for infecting corresponding cells [35]. Especially in higher species, due to the development of generative cells and the mechanism of sexual reproduction, the diffusion of free genome including endogenous virus of cells has been greatly restricted, to experience the further selective elimination. As in human, the existence of the isolation of the placental barrier (except of the viral sequences that are integrated in the cell genome [36,37]), there is really no endogenous virus confirmed. However, since the endogenous virus group is the primitive source of modern viruses and its concealment with the host due to their compatibility, recently scientists has paid great attention to finding the typical representative of the endogenous virus with significance to human beings [38-40].
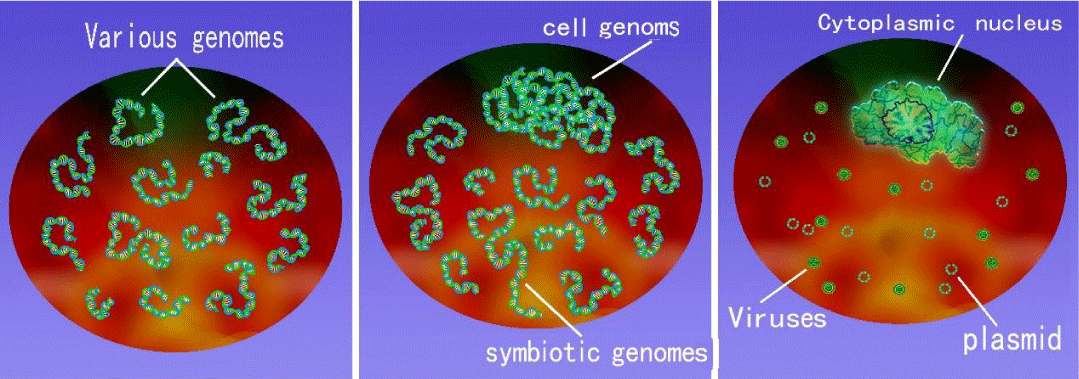 Figure 1: Genome evolution:
Genome stage - Cell genome period - Genome differentiation phase (from left to right).
Figure 1: Genome evolution:
Genome stage - Cell genome period - Genome differentiation phase (from left to right).
The trans-species infection and cyto-pathogenic virulence, group of exogenous virus
Since the tiny virus has very fast copy rate in replication and lacks of the replication error correction mechanism, consequently, the copy error for virus is relatively high. Even in one host cell, the virus really exists in a group state with a variety of nucleic acid sequences in replication and proliferation, in other words, it is in the form of quasi-species and has a quasi-species spectrum [41]. The virus sequence analysis are often the representation of the dominant strains in the quasi-species spectrum. Certain base changes of the virus sequence in replication can lead to significant changes in the performance of the virus, it is also named as gene mutation. When the surface antigen changes in a virus by mutation, it can be combined with ligands or receptor to get into cells of other species, this invasion process is named trans-species infection. For example, the avian influenza virus and mouse Hantan virus invades human beings [42,43]. Dissemination by mutation and trans-species infection are the important mechanisms for expansion and evolution of viruses. Its occurrence depends on the random variation of surface antigen of virus, the contact density of inter-species and genetic relationship of inter-species in the evolution history [44]. When the virus enters the cell, the process of replication and correspondent products affect cell metabolism; they might lead to the cell death or disease; this is defined as viral cytotoxicity [45]. Cytotoxicity is a common feature of exogenous virus infection, and the intensity of virulence depends on the number of invading viruses and the degree of incompatibility with the cells. Due to incompatibility, the exogenous virus and hosts do not coexist peacefully, and the hosts cannot carry viruses to be prevalent within the group, the mortality rate of this kind of infection is high if without effective antiviral drugs, or if subsequent immune response does not remove the virus effectively, for example, human beings have been invaded by avian influenza, rabies virus or hantan virus infection etc., [46-48]. Viral cytotoxicity occurs due to its incompatibility with the cell. In some cases, the cytotoxic phenomenon can also occur by trans-cell infection between different types of cells in one body with multicellular organisms, which may lead disease progression with high mortality. This mechanism can be applied to the pathogenesis of some viral infections. For example, HIV can be hidden in macrophages without cell lesions for a long time but can cause CD4 cells damage by trans-cell infection [49-51], and the similar way occurs with the human influenza virus in brain neurons [52].
Trans-species evolution and Permanent resident virus group
Due to characteristics of virus survival in quasi-species, the spectrums of virus within a trans-species invasion is gradually expanded with replication, which is also shown in a spectrum of differences in virulence [53]. Cell deaths caused by strains of high virulent are difficult to avoid. While low virulence strains may be relatively easier to deal with resulting temporary extension. With the advantage of survival, virus of quasi-species gradually shifts towards low toxicity, the adapted strains might be able to spread among the main flock in the new host, this phenomenon is an important feature of virus survival, called “Passage attenuation of toxicity” [54]. Finally, the avirulent strain is eventually evolved into a subclass of homologous virus with a new species as a long-term host. This kind of trans-species evolution is the main path and the universal law of virus development. For example, the influenza virus, through the evolution of trans-species, has been differentiated into various subtypes’ species respectively in human beings, birds, pigs, dogs and others [55,56]. By sequence analysis of known viruses, most of the viruses associated with human beings have the evolution trace of exogenous viruses, the root of virus could be found in the surrounding species, this is a gradual formation of evolutionary history [57]. Although homologous flus of bird, pig, dog and human show the similar sequences due to trans-species evolution, but each has its own long-term host of the compatibilities, they represent completely different Infective and epidemic regularity in different hosts. Because of the dependence of the virus on their host, it has not been found in nature that one subtype of virus can be prevalent in two species in the same period. Viral nucleic acid sequence analysis is the only one suitable for the study of the evolution of virus and its historical origin, but to determine the infectivity, virulence and relevant epidemiological information of the virus for certain species is very difficult. The trans-species evolution of virus is gradual and probabilistic If the virus strain is poorly compatible with the new host the conflicts between virus and cells could cause the hosts to die; the virus in this case is difficult to survive, trans-species evolution of virus may be naturally eradicated. For example, rabies virus and hantavirus can infect humans, but they fail to get evolved with human as a long-term host.
The subtypes of virus evolved by trans-species are similar to those of endogenous virus group in the sense that they have no cytotoxicity to their new host as long-term host. However, the process of trans-species infection and evolution of virus occur in the level of somatic cells, virus still requires a process of external invasion for its spreading and development generation after generation so it should be called the permanent resident virus by distinction from endogenous virus group originated with cell. The reproductive mechanism of species, especially the mechanism of sexual reproduction, is not only one of the evolutionary mechanisms for exclusion of inferior genes, but also one to exclude free genomes like virus. Even if the endogenous virus does not cross the reproductive barrier during evolution, the infection of new individuals also requires a re-invasion process similar to that of permanent resident viruses. Mutation, trans-species infection, “Passage attenuation of toxicity” and trans-species evolution are the important mechanisms for study the origin of virus and viral diseases. It is an important law of the development of the virus evolution from exogenous virus group to permanent resident virus group.
Immune system and Immuno-pathologic virulence
The cross species infection can lead cells to cytopathic or death, in theory, viral cytotoxicity is likely to destroy cell species. Therefore, the virus presence promotes host to develop the mechanism against the invasion of foreign nucleic acids (viral genome). Modern research on the understanding of the interferon system and CRISPS-CAS system are the mechanisms for the removal of foreign nucleic acids at the cellular level [58,59]. With the development of multicellular species, it has prompted a more complex and effective mechanism to resist and eliminate the invasion of foreign microorganisms - the occurrence, evolution and perfection of the immune system and immunity [60]. Therefore, the development and evolution of the virus across species have a new restriction mechanism. The immune system is a tissue system of high-grade species for resisting invasion of microorganisms including viruses and maintaining the stability of the internal environment. Virus invasion can stimulate host immune response, the functional effects of response can clear the virus and infected cells, and produce the immune-protection against same virus invading again [61]. However, the immune pathological phenomenon presented with the host cell damage and tissue inflammatory responses associated with immune clearance reactions is known as immune toxicity [62]. When the immune response is timely and appropriate, the occurrence of immune pathology can be manifested as a clinical recessive process, but otherwise, could be showing a dominant and even more severe disease process on host if with an excessive or inadequate immune response. Since the occurrence of immune pathological effect is closely related with immune response cycle, so that the disease process is very regular. From the occurrence and mechanism of immunity, Immune function is developed in multicellular organisms on the basis of self or non-self-recognition between cells, this recognition and identification are based on the three-dimensional structure information of biological molecules rather than nucleic acid sequence information. The function process of immune effect cells and their immune molecules have to be carried out on the molecular recognition. They do not have the mechanism to enter cells to recognize and remove nucleic acids of virus. The immune function can clean out free viral particles or infected cells by identifying viral protein expressed on the envelops or membranes. For the group of permanent resident virus with no cytotoxicity, normal immune response can clear free viral particles or the infected cells with viral antigen expressed on membranes, and effectively inhibit the spreading of virus and the occurrence of viral diseases, but is unable to enter the infected cells to recognize and eradicate any intracellular protein antigens or nucleotides of the viruses. Consequently, the virus of resident virus group can be residual in cells and survive with limited proliferation for a long time under the immune pressure, meanwhile to keep the immune system stimulated for maintaining the immune tension. The dynamic balance between the residual virus and the immune tension, and the long term existence of viruses in the presence of immune responses is the normal behavior and ordinary state of permanent resident virus group [63-66]. It is really the basis for keeping the host’s lifelong immunity after infection with virus of permanent resident group, and it is also the material basis and principle that the live vaccine of virus for immune persistence is better than the pure antigenic vaccine [67-70], new individuals need to acquire this lifelong features of immune protection through initial natural infection or live vaccination procedures [70].
Permanent resident virus and Immuno-pathology mechanism
The symbiotic mechanism between the permanent resident viruses and the host has been taking place a great change under the development of the immune system, though the virus with no cytotoxic can be parasitic in a host for a long-term, but immuno-toxicity induced by the immune response of initial infection can lead to immune-pathogenesis in a host, which are the main cause and pathogenesis of infective diseases with permanent resident virus [71,72]. Due to the regularity of the immune reaction response cycle, there is a latent period between the initial attack of virus and disease manifestation induced by Immune toxicity in clinic [73]. Before the immune effect can take place, virus can freely duplicate in a host due to the absence of immune pressure, the infected person becomes an asymptomatic carrier for virus spreading in a sensitive group, and to cause widespread infection and large-scale epidemic prior to the onset of the apparent clinical manifestation in population. In addition to isolating sources of infection, the stabilization of the epidemic depends mainly on the protection of group immunity formed by the universal infection or applications of vaccine [74]. Now, all the well-known epidemic viral diseases affecting human belong to the group of permanent resident virus, such as measles virus, small pox virus, polio virus, varicella, mumps, rubella, HAV, HBV, Flu, HIV, and HPV, etc. and the recent known viruses such as SARS, Ebola, MERS, Zika and so on according their behaviors of epidemic and pathogenesis. The immune-pathology is the common pathogenic mechanism for permanent resident virus group. The difference in the clinical manifestations of the disease is only due to differences in the route of infection and the invasion of cellular tissue, as well as the intensity of the host’s immune response. It is well known that there are two mechanisms that can lead to produce the immune toxicity by the permanent resident virus group. The first is through the initial infection, the permanent resident virus group is retained in persons and to be the important composition of the virus micro ecology in the hosts. It is also a latent source of virus under the dynamic balance between the virus proliferation and immune pressure under the herd immunity protection; it will not lead to epidemic, and this is the normal existence state of this virus group in nature. However, new individuals in the population are therefore difficult to avoid the initial infection, which is the main virus group affecting newborn babies with an average of 5-6 overt infections per year, and a greater base of latent infections. The second is that the permanent resident virus produce new immune toxicity caused by the antigen mutation of virus, the residual virus can proliferate and spread again because of the original herd immune protection is no longer effective to result in a new epidemic situation. It is also the main factor that causes the occurrence of viral epidemic in adults; the occurrence is an abnormal state of permanent resident virus with the antigenic mutation. Finally, it is necessary to form a new herd immune protection to create a new balance for returning to normal state as before, such as the periodic epidemic of human influenza virus. The antigenic mutation of viral breakthrough immune protection under the immune pressure is the driving force of the virus for survival and development with natural law, and no way to avoid it. Sometimes, the resurgence of some endogenous virus or permanent resident virus with immune failure is interior factor for some collapse infection in a clinical environment. It is of great theoretical significance to understand the relationship between the permanent resident virus group and the immunity for understanding the occurrence of viral diseases and how to prevent them. Viral diseases due to immune pathogenesis have acute periodic features. Chronic viral diseases such as HIV / AIDS have nothing to do with this mechanism, as AIDS are mostly caused by cytotoxic effect of cross infection from macrophage cells to CD4 cells leading to the failure with immune toxicity. Chronic hepatitis B is not directly associated with viral infection, but with viral core protein mutations and metastasis [75].
Emerging virus
The concept and definition of the new virus has not yet been defined. In the modern world where we live now, there is no way to reproduce the origin of genes to produce a new viral genome when they first appeared, the new virus must come from the existing virus world, which has only been newly discovered or newly identified. For this work, we just call them the new discovered virus. The discovery was identified in two ways, first through clinical disease such as SARSMERS, Abola, Zika etc, the other way by laboratory nucleic acid screening tests, for example, the recently reported that 1445 new RNA viruses have been found by China CDC [76], such findings are not yet positively linked to disease, and may have potential clinical and research implications. Otherwise, the viruses mutate to produce new trans-species infections as the avian influenza to attack human. It is like a new virus for human, essentially a variant of avian flu. The third situation, one of permanent resident virus acquires a new immunogen because of its mutation, which breaks the previous group immune protection, and produce epidemic diseases with new immune toxicity, it is also like a new virus for original host, for example in the new epidemic strain of influenza virus and the SARS virus derived from human coronavirus [77]. This kind of new strains of the virus is created on the mutation of existence virus, none of them are really original ones, so these viruses should be called emerging virus. Meanwhile In the course of a viral disease, the discovery of similar viruses in other species can only indicates the historical origin of viral evolution, and not to be confirmed of a direct source of infection.
The classification of Human Virology and the prevention and treatment of viral diseases
The classification of the international taxonomy of viruses is based on the structure and sequence of viruses, independent of the hosts [78], which is conducive to the laboratory research. However, there is little clinical significance with virus studies without mention the host because viruses have never existed independently in nature without a host. Laboratory nucleic acid sequence analysis is not able to determine the degree of infectivity and virulence of the virus for any species. Any findings in the Lab research should be further validated in clinical research. Based on the virus’s absolute dependence on its host cells, human virology studies need to target viruses closely related to human health and disease. Therefore, Human Virology should be clearly defined in terms of a human host as well. The classification should be based on cellular compatibility (cytotoxicity) and immune compatibility (immune toxicity) between viruses and their human hosts; it is beneficial to the study of epidemiology, pathogenesis and prevention and treatment of human viral diseases. In recent years, the endogenous virus group with concealment has gradually been recognized by scientists. It is difficult to determine whether the particle is a subsidiary or an independent endogenous virus before its biological properties are explored. Now biologists are paying serious attention to the endogenous virus such as understanding if the bacterial plasmid is the best gene carrier. Due to the double virulent pathogenic role with cytotoxicity and subsequent immune toxicity, the clinical manifestation is often more serious in infection with exogenous virus group, but the infections are scattered in individuals; they are not likely to occur in a large-scale epidemic in human because human beings cannot be virus carriers as sources of infections. For instance, when the avian influenza virus is prevalent in poultry, but infections with human beings occur only sporadically. Which can be effectively stopped by isolation the source of infections. The treatment for these diseases is to inhibit the proliferation of the virus to reduce cell toxicity, to sustain the life until the immune clearance function and body rehabilitation are initiated. Vaccine prevention is only suitable for a small number of susceptible people in some special districts. Viruses belong to the permanent resident virus group are the most closely related to human health and the prevalence of viral diseases in human beings. For example, when the outbreak of Abola in West Africa occurred, according to the epidemic and the law of onset, it was easy to determine that the infective sources were those from the permanent resident virus group, the pathogenic factor was immune toxicity. Since the tissue attacked and damaged occurred dominantly to the vascular endothelial cells, so that manifestation was mainly the multiple organs hemorrhagic lesions of clinical critical illness. In this case, properly inhibition of immune response to decrease of the intensity of immune pathology can reduce the mortality rate. Isolation and reduction of population flow can prevent the expansion of the affected areas without the need to find sources of infection from other species. In the case of overt infection, the infected area covers a larger base of population. The epidemic can naturally come to the end when the population immunization is established. It is wrong and dangerous for an outside susceptible person to enter the epidemic area. An overwhelming flow of overt or covert infection may cause future epidemic areas to expand.
Vaccine research and permanent resident virus
With the biological evolution, the permanent resident virus group in human coexists with human beings as a long-term host, the source of infection just comes from the bodies with temporary imbalance between residual virus and immune-pressure in human population. Each individual human being is difficult to avoid of infections in his/her whole life. The immune toxicity of primary infection excitation is the main pathological mechanism of this kind of viral disease, and the correspondent vaccine is the most effective measure to prevent the occurrence of these kind of viral diseases [79]. The mechanism of the effects of the vaccine application is to provide an artificial infection of a viral agent with the immune toxicity reduced or controlled to an appropriate safe range, the resulting immune protective response can resist subsequent natural infection of the correspondent virus. Attenuated live vaccine infections are involved in the composition of host microecology to maintain the lifelong immunity [80], though the dead vaccine or the viral antigen vaccine need to be used for some times to maintain immune protection as poliovirus [81]. The infant immunization program is aimed at the permanent resident virus group in a normal state with a definite and stable immunogenicity of the virus strain. Vaccines have played a clear role in the prevention of such viral diseases [79]. The vaccines and its protective effects are mainly related to prevent the development of related diseases but they are unable to eliminate virus at all. For example, measles and polio have basically disappeared due to the use of vaccines, but under the immune pressure, this residue virus of infection or vaccinate can still persist in population with the possibility of recovery toxicity for a long time [82], so the infant immunization program should be maintained for a long period [83,84]. The misconception that eradication of human smallpox is due to the fact that the relevant virus is eliminated. The reality is that the virus is not really eliminated but substituted by the vaccinia [85]. The preventive effect of vaccine on viral diseases for adults is not satisfactory. This is because that the virus is being at the abnormal state, there is a new immune toxicity happened in permanent resident virus which breaks through the previous immune protection with immunogenic variation by virus mutation for disease prevalent in adults. The immunogen is different from the normal vaccine used for infants for the scheduled immunization. The application of vaccines requires rapid identification or prediction of new immunogen of the prevalent strains of the virus. For example, in theory each Flu epidemic occurred in adult requires the re-verification and validation of the immunogenicity of vaccine prophylaxis. Similar to the development and applications of influenza vaccines, the development of SARS and Abola vaccine also need to be further explored [86,87]. At present, the prevalence of these viral diseases is all naturally subsided without vaccines. The method of isolation of infectious source plays an important role in these epidemics, so the effect of vaccine use needs to be evaluated in future. The use of vaccines is to prevent viral diseases associated with immune pathogenesis, it cannot exceed the functional range of the immune system, HIV also belongs to the human resident virus group, because the virus can be hidden and spread along the host blood T- cells [88], so the current thinking and methods of vaccine development are deficient [89]. The decline of the infection rate in recent years is due to the role of isolation, not due to vaccine. In theory, vaccines also cannot prevent the occurrence of AIDS, it is because that its pathogeneses prefer to involve in cytotoxic by trans-infection from macrophage to CD4 cells than with immune pathogenesis. Hepatitis B virus (HBV) has become an endogenous virus in chronic HBV carrier with immune tolerance of specific antigen, HBV can exist in hepatocyte and circulatory system freely, so chronic hepatitis B is no longer an infectious disease in the traditional sense. Therefore, we believe that the current global wide application of vaccine cannot prevent chronic HBV infection [90]. The failure of hepatitis B vaccine for treatment is understandable [91], if the vaccine were successful, the consequences of stimulating immune toxicity would be clinically difficult to estimate. Another disease closely related to the permanent resident virus is the virus associated with tumor. In theory, the interference of human genome or the activation of tumor gene by viral genomes reside in cells is one of the important mechanisms of tumorigenesis, as HBV for hepatocellular carcinoma [92] and HPV for cervical cancer [93,94]. Vaccines can inhibit viral diseases associated with immune pathogenesis, but cannot eliminate the virus with tumorigenic factors in cells completely. In our opinion, the anti-tumor vaccine should target toward tumor specific antigen.
The Transmigration of Viruses
A harmonious coexistence of viruses with long-term hosts is the normal state in nature. Disease is a state of disorder caused by mutations that cause viruses to conflict with their hosts, this disorder state would return to the normal state through the passage of attenuated cytotoxicity or reconstruction of immune balance between virus and the host. Trans-species infection, passage attenuated cytotoxicity, trans-species evolution and cross reproductive barrier are the natural laws of virus development and evolution. We believe that the viruses of human virology should be classified according to their compatibility characteristics with human being hosts. By doing that, it will be beneficial to basic and clinical study of viruses and viral diseases with human beings. Based on above classification and the principles of evolution, the viral groups described above can evolve from each other relative to the host. For instance, HIV, such as simian virus, has evolved from an exogenous virus group into a permanent resident virus group as a result of changes in cell compatibility with human [95]. HBV as a permanent resident virus of human has improved immune compatibility with mother to child transmission, and has evolved into an endogenous virus group in some populations [96]. The occurrence of emerging virus is not possible to repeat the origin of life in the ancient times. Often, they must have derived from one type of the original virus. When a virus of permanent resident viruses infects another species as trans-infection, a new round of attenuated passages would happen. In the species with immune system, the variation of immunogen of a permanent resident virus will lead to a new round of immune compatibility rebuilding process to achieve a new immune balance. With the above classification of Human Virology, it is beneficial to identify each newly occurring virus or viral disease quickly and clearly in order to take corresponding prevention measures. As recent reported Zika viruses [97], according to its epidemic characteristics, such as long-term existence in virus carrier without cytotoxicity and immune toxicity, and its concealment and particularity of disease, the Zika virus should belong to the endogenous virus. The pathogenic mechanism may be related to the interference of cell genome in related cells over a long period of time (Figure 2). The coexistence of viruses and hosts and their evolution are naturally endowed, these endogenous or permanent resident virus are the best vehicle for gene therapy [98]. The application of nucleoside antiviral drugs will change the natural evolution process of the virus [99]. Human beings are aware and gradually pay attention to its possible consequences; we must conform to this natural internal force of virus and use viruses to serve the people.
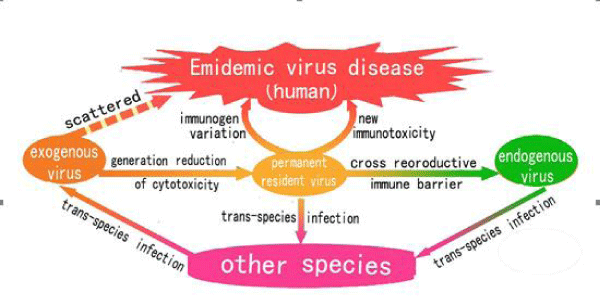 Figure 2: Schematic diagram of virus evolution.
Figure 2: Schematic diagram of virus evolution.
Acknowledgement
This work is supported by National Natural Science Foundation of China (Grant Number: 81127901).
References
- Li Z, Shao Y, Liu C, Liu D, Guo D, Qiu Z, et al. Isolation and pathogenicity of the mammalian orthoreovirus MPC/04 from masked civet cats. Infect Genet Evol. 2015; 36: 55-61. https://goo.gl/NTU4mG
- Lau SK, Feng Y, Chen H, Luk HK, Yang WH, Li KS, et al. Severe Acute Respiratory Syndrome (SARS) coronavirus ORF8 protein is acquired from SARS-related coronavirus from greater horseshoe bats through recombination. J Virol. 2015; 89: 10532-10547. https://goo.gl/8KfoAg
- Reed PE, Mulangu S, Cameron KN, Ondzie AU, Joly D, Bermejo M, et al. A new approach for monitoring Ebola virus in wild great apes. PLoS Negl Trop Dis. 2014; 8: 3143. https://goo.gl/J3JChc
- Groseth A, Feldmann H, Strong JE. The ecology of Ebola virus. Trends Microbiol. 2007; 15: 408-416. https://goo.gl/sWxzZu
- Greenwood B. The contribution of vaccination to global health: past, present and future. Philos Trans R Soc Lond B Biol Sci. 2014; 369: 20130433. https://goo.gl/BwMNXF
- Henderson DA. The eradication of smallpox: an overview of the past, present, and future. Vaccine. 2011; 29: 7-9. https://goo.gl/WC4gSx
- McMichael A, Picker LJ, Moore JP, Burton DR. Another HIV vaccine failure: where to next? Nat Med. 2013; 19: 1576-1577. https://goo.gl/i2oF9E
- Meireles LC, Marinho RT, Van Damme P. Three decades of hepatitis B control with vaccination. World J Hepatol. 2015; 7: 2127-2132. https://goo.gl/3q5SFp
- Zuckerman JN, Sabin C, Craig FM, Williams A, Zuckerman AJ. Immune response to a new hepatitis B vaccine in healthcare workers who had not responded to standard vaccine: randomised double blind dose-response study. BMJ. 1997; 314: 329-333. https://goo.gl/AVWdr8
- Debing Y, Neyts J, Delang L. The future of antivirals: broad-spectrum inhibitors. Curr Opin Infect Dis. 2015; 28: 596-602. https://goo.gl/WTxuUK
- Ackermann M, Engels M, Fraefel C, Metzler A, Schwyzer M, Suter M, et al. Herpesviruses: balance in power and powers imbalanced. Vet Microbiol. 2002; 86: 175-181. https://goo.gl/qtWXQj
- McKee SJ, Bergot AS, Leggatt GR. Recent progress in vaccination against human papillomavirus-mediated cervical cancer. Rev Med Virol. 2015; 25: 54-71. https://goo.gl/7GiM7n
- Lincoln T, Joyce G. Self-sustained replication of an RNA enzyme. Science. 2009; 323: 1229-1232. https://goo.gl/yhyU6J
- Gebicki JM, Hicks M. Ufasomes are stable particles surrounded by unsaturated fatty acid membranes. Nature. 1973; 243: 232-234. https://goo.gl/ES7XZo
- Deamer D, Dworkin JP, Sandford SA, Bernstein MP, Allamandola LJ. The first cell membranes. Astrobiology. 2002; 2: 371-381. https://goo.gl/zV6DCD
- Gold L, Polisky B, Uhlenbeck O, Yarus M. Diversity of oligonucleotide functions. Annu Rev Biochem. 1995; 64: 763-797. https://goo.gl/v8crjm
- Osborne SE, Ellington AD. Nucleic acid selection and the challenge of combinatorial chemistry. Chem Rev. 1997; 97: 349-370. https://goo.gl/JeFTC9
- Pressman A, Blanco C, Chen I. The RNA world as a model system to study the origin of life. Curr Biol. 2015; 25: 953-963. https://goo.gl/ts38cY
- Root-Bernstein M, Root-Bernstein R. The ribosome as a missing link in the evolution of life. J Theor Biol. 2015; 367: 130-158. https://goo.gl/8uYmJS
- Zhu YY, Machleder EM, Chenchik A, Li R, Siebert PD. Reverse transcriptase template switching: a SMART approach for full-length cDNA library construction. Biotechniques. 2001; 30: 892-897. https://goo.gl/sWeksW
- Kibria MG, Chowdhury FA, Trudeau ML, Guo H, Mi Z. Dye-sensitized InGaN nanowire arrays for efficient hydrogen production under visible light irradiation. Nanotechnology. 2015; 26: 285401. https://goo.gl/6g3YmK
- Dodekatos G, Schünemann S, Tüysüz H. Surface plasmon-assisted solar energy conversion. Top Curr Chem. 2016; 371: 215-252. https://goo.gl/GKpg8a
- Mangels D, Kruip J, Berry S, Rögner M, Boekema EJ, Koenig F. Photosystem I from the unusual cyanobacterium Gloeobacter violaceus. Photosynth Res. 2002; 72: 307-319. https://goo.gl/ji24xe
- Noller H.F. Evolution of protein synthesis from an RNA world. Cold Spring Harb Perspect Biol. 2012; 4: 003681. https://goo.gl/zKLBNL
- Foster SD, Oram SH, Wilson NK, Gottgens B. From genes to cells to tissues modelling the haematopoietic system. Mol Biosyst. 2009; 5: 1413-1420. https://goo.gl/ybMng5
- Kaniak-Golik A, Skoneczna A. Mitochondria-nucleus network for genome stability. Free Radic Biol Med. 2015; 82: 73-104. https://goo.gl/gww4kC
- Martin W, Rujan T, Richly E, Hansen A, Cornelsen S, Lins T, et al. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc Natl Acad Sci U S A. 2002; 99: 12246-12251. https://goo.gl/xJ4XCh
- Flores R, Gago-Zachert S, Serra P, Sanjuan R, Elena SF. Viroids: survivors from the RNA world? Annu Rev Microbiol. 2014; 68: 395-414. https://goo.gl/ftDaum
- Carattoli A. Plasmids and the spread of resistance. Int J Med Microbiol. 2013; 303: 298-304. https://goo.gl/y854wJ
- Tazzyman SJ, Bonhoeffer S. Why there are no essential genes on plasmids. Mol Biol Evol. 2015; 32: 3079-3088. https://goo.gl/z3t6GR
- Mizokami M, Orito E, Ohba K, Ikeo K, Lau JY, Gojobori T. Constrained evolution with respect to gene overlap of hepatitis B virus. J Mol Evol. 1997; 44: 83-90. https://goo.gl/kjtwru
- Crick F.H. The origin of the genetic code. J Mol Biol. 1968; 38: 367-379. https://goo.gl/fdqBkE
- Butler D. Flu genomes trace H7N9 evolution. Nature. 2015; 519: 137. https://goo.gl/XVuAe8
- Flowers EM, Bachvaroff TR, Warg JV, Neill JD, Killian ML, Vinagre AS, et al. Genome sequence analysis of CsRV1: A pathogenic reovirus that infects the blue crab callinectes sapidus across its trans-hemispheric range. Front Microbiol. 2016; 7: 126. https://goo.gl/obp9er
- Flint SJ, Racaniello VR, Enquist LW, Skalka AM. Chapter 2: Infection of populations. Volume II Pathogenesis and control; In Principles of virology. ASM Press American Society for Microbiology Washington DC. 2009; 28-51. https://goo.gl/pj2Hu1
- Nowrouzi A, Glimm H, von Kalle C, Schmidt M. Retroviral vectors: post entry events and genomic alterations. Viruses. 2011; 3: 429-455. https://goo.gl/4KrGoM
- Hughes JF, Coffin JM. Evidence for genomic rearrangements mediated by human endogenous retroviruses during primate evolution. Nat. Genet. 29, 487−489. Nat Genet. 2001; 29: 487-489. https://goo.gl/a4VqQP
- Denner J, Petersen B, Niemann H. Tolerance and immune response to the porcine endogenous retrovirus in German landrace pigs immunised with viral proteins. Virus Res. 2015; 208: 39-43. https://goo.gl/g3dbYo
- Li W, Lee MH, Henderson L, Tyagi R, Bachani M, Steiner J, et al. Human endogenous retrovirus-K contributes to motor neuron disease. Sci Transl Med. 2015; 7: 307. https://goo.gl/jfWYaB
- Contreras-Galindo R, Kaplan MH, Dube D, Gonzalez-Hernandez MJ, Chan S, Meng F, et al. Human endogenous retrovirus type K (HERV-K) particles package and transmit HERV-K-related sequences. J Virol. 2015 Jul; 89: 7187-7201. https://goo.gl/s9vpPW
- Domingo E. Quasispecies theory in virology. Journal of Virology. 2002; 76: 463-465. https://goo.gl/oXwfzS
- Velumani S, Du Q, Fenner BJ, Prabakaran M, Wee LC, Nuo LY, et al., Development of an antigen-capture ELISA for detection of H7 subtype avian influenza from experimentally infected chickens. J Virol Methods. 2008; 147: 219-225. https://goo.gl/mcx64T
- Jay Hooper, Connie S. Schmaljohn. Hantavirus. Springer Index of Viruses. 2002; 150-156. https://goo.gl/NmrMV5
- Wagnerova M, Chalupkova A, Hrabovska Z, Ancicova L, Mistrikova J. Possible role of different animal species in maintenance and spread of murine gammaherpesvirus 68 in the nature. Acta Virol. 2015; 59: 14-19. https://goo.gl/5BbW5a
- Mahnel H. Cytotoxicity of Virus. Cytotoxicity of Virus. 1968.
- Ewald SJ, Kapczynski DR, Livant EJ, Suarez DL, Ralph J, McLeod S, et al. Association of Mx1 Asn631 variant alleles with reductions in morbidity, early mortality, viral shedding, and cytokine responses in chickens infected with a highly pathogenic avian influenza virus. Immunogenetics. 2011; 63: 363-375. https://goo.gl/6nxLjw
- CC. Human rabies Virginia. MMWR Morb Mortal Wkly Rep. 2010; 59: 1236-1238. https://goo.gl/9jW63g
- Altun DU, Öktem IMA, Ünal B. Hantavirus infections and prevention of Hantaviruses. TAF Prev Med Bull. 2011; 10: 373-378. https://goo.gl/zBGPNf
- Sterjovski J, Churchill MJ, Ellett A, Gray LR, Roche MJ, Dunfee RL, et al. Asn 362 in gp120 contributes to enhanced fusogenicity by CCR5-restricted HIV-1 envelope glycoprotein variants from patients with AIDS. Retrovirology. 2007; 4: 89. https://goo.gl/umgvWF
- Xiao X, Wu L, Stantchev TS, Feng YR, Ugolini S, Chen H, et al. Constitutive cell surface association between CD4 and CCR5. Proc Natl Acad Sci U S A. 1999; 96: 7496-7501. https://goo.gl/tdFWXv
- Endres MJ, Clapham PR, Marsh M, Ahuja M, Turner JD, McKnight A, et al. CD4-independent infection by HIV-2 is mediated by Fusin/CXCR4. Cell. 1996; 87: 745-756. https://goo.gl/6KcAcb
- Wisniewski HM, Brown HR, Thormar H. Pathogenesis of viral encephalitis: Demonstration of viral antigen(s) in the brain endothelium. Acta Neuropathol. 1983; 60: 107-112. https://goo.gl/PeK3Tu
- Sanz-Ramos M, Rodríguez-Calvo T, Sevilla N. Mutagenesis-mediated decrease of pathogenicity as a feature of the mutant spectrum of a viral population. PLOS One. 2012; 7: 39941. https://goo.gl/4NRFgU
- Lee CA, August A, Arnold JJ, Cameron CE. Polymerase mechanism-based method of viral attenuation. Methods Mol Biol. 2016; 1349: 83-104. https://goo.gl/bSdwQV
- Ma S, Downard KM, Wong JW. FluClass: A novel algorithm and approach to score and visualize the phylogeny of the influenza virus using mass spectrometry. Anal Chim Acta. 2015; 895: 54-61. https://goo.gl/Dca5h3
- Li Q, Wang X, Sun Z, Hu J, Gao Z, Hao X, et al. Adaptive mutations in PB2 gene contribute to the high virulence of a natural reassortant H5N2 avian influenza virus in mice. Virus Res. 2015; 210: 255-263. https://goo.gl/cYbRz5
- Eigen M. The origin of genetic information: viruses as models. Gene. 1993; 135: 37-47. https://goo.gl/Vej6BA
- Qi Z, Nie P, Secombes CJ, Zou J. Intron-containing type I and type III IFN coexist in amphibians: refuting the concept that a retroposition event gave rise to type I IFNs. J Immunol. 2010; 9: 5038-5046. https://goo.gl/sTpmKL
- Makarova KS, Wolf YI, Alkhnbashi OS, Costa F, Shah SA, Saunders SJ, et al. An updated evolutionary classification of CRISPR-Cas systems. Nat Rev Microbiol. 2015; 13: 722-736. https://goo.gl/6BjQ2t
- Jackson JA. Immunology in wild nonmodel rodents: an ecological context for studies of health and disease. Parasite Immunol. 2015; 37: 220-232. https://goo.gl/zKJu3v
- Barker E. Immunity to virus. Immunology, Infection, and Immunity. Gerald B. Pier (ASM Press. Washington D.C). 2004; 463-467.
- Rooney AA, Luebke RW, Selgrade MK, Germolec DR. Immunotoxicology and its application in risk assessment. EXS. 2012; 101: 251-287. https://goo.gl/5WU3Rc
- Xu L, Wei Y, Chen T, Lu J, Zhu CL, Ni Z, et al. Occult HBV infection in anti-HBs-positive young adults after neonatal HB vaccination. Vaccine. 2010; 28: 5986-5992. https://goo.gl/75U6Z5
- Zerbini A, Pilli M, Boni C, Fisicaro P, Penna A, Di Vincenzo P, et al., The characteristics of the cell-mediated immune response identify different profiles of occult hepatitis B virus infection. Gastroenterology. 2008; 134: 1470-1481. https://goo.gl/cX7JPS
- Lellouch Tubiana A, Fohlen M, Robain O, Rozenberg F. Immunocytochemical characterization of long-term persistent immune activation in human brain after herpes simplex encephalitis. Neuropathol Appl Neurobiol. 2000; 26: 285-294. https://goo.gl/d9nGyN
- Brockman MA, Knipe DM. Herpes simplex virus vectors elicit durable immune responses in the presence of preexisting host immunity. J Virol. 2002; 76: 3678-3687. https://goo.gl/JeVf2P
- Bandyopadhyay AS, Garon J, Seib K, Orenstein WA. Polio vaccination: past, present and future. Future Microbiol. 2015; 10: 791-808. https://goo.gl/DLq3tA
- Okayasu H, Sein C, Hamidi A, Bakker WA, Sutter RW. Development of inactivated poliovirus vaccine from Sabin strains: A progress report. Biologicals. 2016; 44: 581-587. https://goo.gl/62T7JK
- Abe S, Ota Y, Koike S, Kurata T, Horie H, Nomura T, et al. Neurovirulence test for oral live poliovaccines using poliovirus-sensitive transgenic mice. Virology. 1995; 206: 1075-1083. https://goo.gl/ddrp9a
- Isanaka S, Guindo O, Langendorf C, Matar Seck A, Plikaytis BD, Sayinzoga-Makombe N et al. Efficacy of a low-cost, heat-stable oral rotavirus vaccine in Niger. N Engl J Med. 2017; 376: 1121-1130. https://goo.gl/FwdMn7
- Moriyama T, Guilhot S, Klopchin K, Moss B, Pinkert CA, Palmiter RD, et al. Immunobiology and pathogenesis of hepatocellular injury in hepatitis B virus transgenic mice. Science. 1990; 248: 361-364. https://goo.gl/sPjo1T
- Chisari FV. Hepatitis B virus transgenic mice: Models of viral immunobiology and pathogenesis, in transgenic models of human viral and immunological disease. Curr Top Microbiol Immunol. 1996; 206: 149-173. https://goo.gl/9ite4o
- Hermesh T, Moltedo B, López CB, Moran TM. Buying time-the immune system determinants of the incubation period to respiratory viruses. Viruses. 2010; 2: 2541-2558. https://goo.gl/xttT1o
- Plans-Rubió P. Evaluation of the establishment of herd immunity in the population by means of serological surveys and vaccination coverage. Hum Vaccin Immunother. 2012; 8: 184-188. https://goo.gl/1fT5Zy
- Papatheodoridis GV, Hadziyannis SJ. Diagnosis and management of pre-core mutant chronic hepatitis B. J Viral Hepat. 2001; 8: 311-321. https://goo.gl/b2FVLo
- Shi M, Lin XD, Tian JH, Chen LJ, Chen X, Li CX, et al. Redefining the invertebrate RNA virosphere. Nature. 2016; https://goo.gl/yZ94Nh
- Menachery VD, Yount BL Jr, Debbink K, Agnihothram S, Gralinski LE, Plante JA, et al. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat Med. 2015; 21: 1508-1513. https://goo.gl/ooZydq
- Bos L. International naming of viruses - a digest of recent developments. Arch Virol. 2002; 147: 1471-1477. https://goo.gl/r7cHzx
- Greenwood B. The contribution of vaccination to global health: past, present and future. Philos. Philos Trans R Soc Lond B Biol Sci. 2014; 369: 20130433. https://goo.gl/5TJTsi
- Minor PD. Live attenuated vaccines: Historical successes and current challenges. Virology. 2015; 479-480: 379-92. https://goo.gl/kYMFZu
- SALK JE. Persistence of immunity after administration of formalin-treated poliovirus vaccine. Lancet. 1960; 2: 715-23. https://goo.gl/juBvNK
- Abad CL, Safdar N. The Reemergence of measles. Curr Infect Dis Rep. 2015; 17: 51. https://goo.gl/PmV4kT
- Morgan OW. Following in the footsteps of smallpox: can we achieve the global eradication of measles? BMC Int Health Hum Rights. 2004; 4: 1. https://goo.gl/3FyiDr
- Taniuchi M, Famulare M, Zaman K, Uddin MJ, Upfill-Brown AM, Ahmed T et al. Community transmission of type 2 poliovirus after cessation of trivalent oral polio vaccine in Bangladesh: an open-label cluster-randomised trial and modelling study. Lancet Infect Dis. 2017; 17: 1069-1079. https://goo.gl/oL2Ayy
- Greenberg RN, Kennedy JS. ACAM2000: a newly licensed cell culture-based live vaccinia smallpox vaccine. Expert Opin Investig Drugs. 2008; 17: 555-654. https://goo.gl/2TtpKe
- Berger A, Drosten Ch, Doerr HW, Stürmer M, Preiser W. Severe acute respiratory syndrome (SARS)--paradigm of an emerging viral infection. J Clin Virol. 2004. 29: 13-22. https://goo.gl/vunWoV
- Sharma R, Jangid K, Anuradha. Ebola vaccine: How far are we? J Clin Diagn Res. 2017; 11: 01-04. https://goo.gl/hpZCkf
- Sherer NM, Lehmann MJ, Jimenez-Soto LF, Horensavitz C, Pypaert M, Mothes W. Retroviruses can establish filopodial bridges for efficient cell-to-cell transmission. Nat Cell Biol. 2007; 9: 310-5. https://goo.gl/UoXqQq
- Boasso A, Shearer GM, Clerici M. The hunt for an HIV vaccine: time to rethink recent failures. Lancet. 2008; 371: 1897-1898. https://goo.gl/svFbNJ
- Poovorawan Y, Chongsrisawat V, Theamboonlers A, Leroux-Roels G, Kuriyakose S, Leyssen M, et al. Evidence of protection against clinical and chronic hepatitis B infection 20 years after infant vaccination in a high endemicity region. J Viral Hepat. 2011; 18: 369-375. https://goo.gl/FE8Ppx
- Xu DZ, Wang XY, Shen XL, Gong GZ, Ren H, Guo LM, et al. Results of a phase III clinical trial with an HBsAg-HBIG immunogenic complex therapeutic vaccine for chronic hepatitis B patients: experiences and findings. J Hepatol. 2013; 59: 450-456. https://goo.gl/ipV1jC
- Ringelhan M, O'Connor T, Protzer U, Heikenwalder M, et al. The direct and indirect roles of HBV in liver cancer: prospective markers for HCC screening and potential therapeutic targets. J Pathol. 2015; 235: 355-367. https://goo.gl/qySZsz
- Argyri E, Tsimplaki E, Marketos C, Politis G, Panotopoulou E, et al. Investigating the role of human papillomavirus in lung cancer. Papillomavirus Res. 2017; 3: 7-10. https://goo.gl/34ADS2
- Xue Y, Toh SY, He P, Lim T, Lim D, Pang CL, et al. HPV16-E2 induces prophase arrest and activates the cellular DNA damage response in vitro and in precursor lesions of cervical carcinoma. Oncotarget. 2015; 6: 34979-34991. https://goo.gl/qt1Gvy
- Reid MJC, Switzer WM, Schillaci MA, Klegarth AR, Campbell E, Ragonnet-Cronin M, et al. Bayesian inference reveals ancient origin of simian foamy virus in orangutans. Infect Genet Evol. 2017; 51: 54-66. https://goo.gl/zDPYvv
- Badur S, Lazizi Y, Ugurlu M, Perk Y, Ilter O, Aydinli K, et al. Transplacental passage of hepatitis B virus DNA from hepatitis B e antigen-negative mothers and delayed immune response in newborns. J Infect Dis. 1994; 169: 704-706. https://goo.gl/Mgzhm3
- Rodrigues LC. Microcephaly and Zika virus infection. Lancet. 2016; 387: 2070-2072. https://goo.gl/9U6K9g
- Fountzilas C, Patel S, and Mahalingam D. Review: Oncolytic Virotherapy, updates and future directions. Oncotarget. 2017.
- Mansky LM. Mutagenic outcome of combined antiviral drug treatment during human immunodeficiency virus type 1 replication. Virology. 2003; 307: 116-121. https://goo.gl/azJYcS
Authors submit all Proposals and manuscripts via Electronic Form!