Research Article
Spinach Leaf Extract Ameliorates Multiple Organ Dysfunction Induced by Lipopolysaccharide Exposure in Animal Model of Parkinson's Disease
Monika Kapoor1*, Neha Sharma1* and Bimla Nehru1
1Department of Biophysics, Panjab University, Chandigarh- 160014.
*Address for Correspondence: Dr. Neha Sharma, Department of Biophysics, Punjab University, Chandigarh -160014, India, Tel: +91-8283838703; Email: jaitly_neha@rediffmail.com
Dates: Submitted: 26 July 2017; Approved: 16 August 2017; Published: 18 August 2017
Citation this article: Kapoor M, Sharma N, Nehru B. Spinach Leaf Extract Ameliorates Multiple Organ Dysfunction Induced by Lipopolysaccharide Exposure in Animal Model of Parkinson's Disease. Alzheimers Parkinsons Dis Open Access. 2017;3(1): 001-009.
Copyright: © 2017 Kapoor M, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Keywords: Lipopolysaccharide; Multiple organ dysfunctions; Oxidative burden; Inflammation; Spinach leaf extract
Abstract
Gut microflora is a huge reservoir of Lipopolysaccharide (LPS), an endotoxin which is the main component of external cell wall of gram negative bacteria. A balance exists between the gut microflora and host, the disruption of which leads to many diseases and inflammatory cascades. Various studies have shown that change in gut permeability promote the translocation of microbes into the liver via portal circulation and results in activation of immune system. Along with liver, many other organs are also getting affected including brain and kidney. Literature also revealed that the spinach leaf extract is rich in several active components; including flavonoids and hence exhibit anti-oxidative, anti-proliferative and anti-inflammatory properties in biological system. The present study was aimed to explore the protective potentials of the Spinach Leaf Extract (SLE) (50 mg/kg b.w) against the systemic LPS (5mg/kg b.w) induced toxicity by evaluating several toxicological parameters such as increased oxidative burden in organs as depicted from compromised antioxidant defence system (including Glutathione System, MDA levels, Catalase activity), cytokine levels and blood glucose levels. This was further supported by results from histopathological studies. Accordingly, supplementation with SLE to LPS treated mice showed marked improvement in the above altered parameters. Histopathological study also depicted improved histoarchitecture in liver, kidney and brain after SLE supplementation. Above stated data suggested that treatment with spinach leaf extract protects against LPS induced multiple organ failure by virtue of its antioxidant properties. It also protects by acting on multifactorial pathways simultaneously therefore, its therapeutic potentials should be explored further.
Introduction
Several studies revealed that the number of microbes present in the gut are much more than the total number of cells present in the human body [1]. There are 1012 bacterial cells in each gram of faeces [1]. Microbes in the gut perform important roles in digestion of food, vitamins and cast immunity [2,3]. Alterations in the composition of the microflora or in gut permeability assists in the translocation of microbes into the portal circulation that delivers blood directly to the liver. This microflora is a huge reservoir of Lipo Poly Saccharide (LPS), an endotoxin which is a main component of the external cell wall of Gram-negative bacteria [4].
Liver represents the first line of defence against infection as 70% of the blood supply received from the intestine is via hepatic portal vein. It is equipped with many immune cells (i.e., macrophages i.e Kupfer cells, lymphocytes, natural killer cells, and dendritic cells) to accomplish this function and results in induction of inflammatory cascade via activation of these cells [5]. When activated, kupffer cells secrete a variety of cytokines such as Tumor Necrosis Factor alpha (TNF-a) and several types of pro inflammatory mediators which further induce the production of Reactive Oxygen Species (ROS) such as superoxide anion (O2-) via activation of NADPH oxidise complex [6,7] and Reactive Nitrogen Species (RNS) such as peroxynitrite (ONOO-) by reaction of O2- and NO. The additive inflammatory response finally leads to multiple organ injury that is oxidative burden and toxicity in liver [8]. Acute Renal Failure [9]. And neuro in?ammatory responses in the brain, which was followed by accumulation of Aβ which is involved in the cognitive deficits and memory Impairments [10].
Extensive experimental and observational research realized that life style plays an important role in chronic diseases. Interaction of life style factors with genetic factors leads to the disease process. Edible plants perhaps one of the most important life style components that could modulate the progression of various diseases. Natural plants polyphenols (flavonoids and non-flavonoids) are the most abundant anti-oxidant in the diet and as such are ideal photochemical for neutralizing stress induced by free radicals and inflammation.
NAO (Natural antioxidant) extracted from spinach leaf extract (Spinaciaoleracea) [11] was found to be superior to the well-known antioxidants butylated hydroxytoluene and vitamin E [12]. Potent antioxidant properties including the scavenging of nitric oxide, free reactive oxygen radicals and lipid radicals [13,14]. The anti-inflammatory action of flavonoids is due to prevention of leukotriene production by lipoxygenase in leukocytes [15,16]. NAO contain a mixture of polyphenols that inhibit the lipoxygenase enzyme [12]. Which cause lipid peroxidation. Several studies manifest the neuroprotective efficacy of flavonoids in improving the locomotors activity [17,18]. It was hypothesised that NAO extract from spinach leaf by virtue of its antioxidant as well as anti-inflammatory properties will act upon multi factorial pathways activated by LPS exposure. Therefore, the present study was designed with an aim to explore the protective potentials of spinach leaf extract against LPS induced multiple organ injury.
Materials and Methods
Animals
Healthy male BALB/c mice weighing 30-35 gm were procured from the animal house of the university and were provided standard animal feed and water ad labium throughout the treatment. They were housed in polypropylene cages under hygienic conditions. All procedures were done in accordance with ethical guidelines on the care and use of laboratory animals which were approved by Institutional Animal Ethics Committee (IAEC).
Chemicals
Lipopolysaccharide (LPS) was purchased from Sigma (St. Louis.MO). Spinach Leaf Extract (SLE) was received as a gift sample from BLP Pharmaceutical & Chemical Limited, China.
Experimental Methodology
Animals were divided into four different groups (n = 8-10).
Grouping of animals is following
Group I: Control animals: Animals were given single injection of normal saline (0.9%) (5 ml/kg b.wt, i.p).
Group II: LPS treated animals: The animals in this group were given single injection of LPS (5mg/kg b.wt, i.p).
Group III: LPS+ Spinach leaf extract (SLE) treated animals: Animals in this group were administered with single injection of LPS (5mg/kg b.wt, i.p) and spinach leaf extract (50mg/kg b.wt, p.o) daily for a period of 21 days.
Group IV: Spinach leaf extract treated (SLE) animals: Animals in this group were administered with spinach leaf extract (50mg/kg b.wt, p.o) daily for a period of 21 days.
Behavioral studies
Animals were tested for active avoidance and passive avoidance for cognition and memory.
Passive avoidance test
It was performed to evaluate learning and memory by the method of Miyamoto et al [19]. The apparatus for the test was made of two open chambers and one closed chamber. All of them were interconnected. The animal was positioned in the open chamber; the light from lamp was made to fall on the animal. The time taken by the mice to enter the closed chamber from open light chamber was noted as the acquisition trial time. The shutter was enclosed and the shock of 2.5 mA was given for 3 s. Again after 24 h the animal was put in the lighted chamber and time was noted. The time taken by the animal to enter the enclosed chamber was taken as retention trials. The animal is allowed to stay in the open lighted chamber for a maximum of 300 s.
Active avoidance test
Cognitive behaviour was assessed by the number of times and was performed by the method of Miyamoto et al [19], the animal escapes in the 10 test trial series. To qualify the animal has to avoid at least 8 times out of 10 trials.
Collection of samples
Animals in all groups were sacrificed immediately after performing the behavioural testing. Liver, kidney and brain samples were collected, rinsed in ice-cold normal saline, carefully blotted, weighed and homogenized (10% w/v) in 10 mM of ice cold Phosphate Buffer Saline (PBS, pH 7.4). Homogenate was centrifuged at 1000g for 30 min at 4oC. Pellet was discarded and whole of the supernatant was used.
Biochemical parameters
The following biochemical parameters were done:
Protein estimation: Protein contents in various samples were estimated by the method of Lowry [20].
Lipid Peroxidation: Malondialdehyde levels were assessed in the Post mitochondrial fraction (PMF) in accordance to the method described by Wills [21]. Since malondialdehyde is one of the by-products of lipid per oxidation, levels of malondialdehyde (MDA) defined as an index for determining the extent of lipid per oxidation from breakdown of polyunsaturated fatty acids.
Catalase: Catalase was estimated by using the method as described by Luck [22]. The enzyme catalyses the decomposition of Hydrogen peroxides (H2O2), which is measured by decrease in absorbance, at 240 nm.
Reduced Glutathione (GSH): Reduced glutathione content was estimated according to the method of Ellman [23]. In this method, 5, 5-dithiobis2-nitrobenzoic acid (DTNB) is reduced by -SH groups to form 2-nitro-5-mercaptobenzoic acid per mole of SH. The nitro mercaptobenzoic acid anion released has an intense yellow color and can be used to measure -SH groups at 412 nm.
Determination of Glutathione Peroxidise: Glutathione peroxidise activity was estimated by the method of Paglia and Valentine [24] using H2O2 as substrate. The oxidized glutathione continuously reduced by excess of glutathione reductase and simultaneous oxidation of NADPH monitored at 340 nm.
mRNA Expression studies
mRNA expression analysis by Reverse transcriptase-PCR was performed using one step RT-PCR kit (Invitrogen). Various genes associated with inflammation like TNF-a, IL-1a, IL-1β was also analyzed. Mouse β-actin was used as an internal control. For these studies total RNA isolation was carried out from the mid brain region.
Optimal primer pairs were designed using the software "Gene Runner" or their sequence were obtained from literature and were got synthesized from Sigma Aldrich (USA). Lengths of the primer chosen were approx. 20bp. Gene IL-1β with Gene ID: NM_008361.3, primer sequence (F) 5'-AGCTGCACTGCAGGCTTCGAGATG-3'(R) 5'-GAACTGTGCAGACTCAAACTCCAC-5' and amplification size of 338 bp. Gene TNF-a with Gene ID: NM_013693.2, primer sequence: (F) 5'-ACTGAACTTCGGGGTGATTG-3'(R) 5'-GTGGGTGAGGAGCAGGTAGT-3'and amplification size of 319 bp.PCR products were separated on 2% agarose gels containing ethidium bromide and visualized under ultraviolet light. Densitometry analysis of bands was done by using the Image J software (NIH). The cycle time (Ct) values genes first normalized with β-actins of the same sample, and then the relative differences between control and treatment groups were calculated and expressed as relative change.
Light microscopic examination
To assess histoarchitecture changes if any, small sections of liver, kidney and brain from each of the normal control and different treated animals were taken, washed with ice-cold 0.9% NaCl and were fixed in the buffered formalin (10%) for about 24-48 h. After the fixation, tissues were dehydrated in ascending grade of alcohol, embedded in wax following the standard technique [25]. 5-7 μm thick paraffin sections were cut and then were subjected to haematoxylin-eosin staining of liver and kidney as described by Humanson [26] and cresol violet staining of brain sections as described by Keller [27].
Statistical analysis
For analyzing the data, one way analysis of variance (ANOVA) followed by Newman Keel's test was performed using the statistical software package "SSPS v 11 for windows". The post-hoc comparison of means from different treatment groups were made by the method of Least Significant Difference (LSD). Results corresponding to a p value of 0.05 or less were considered statistically significant.
Results
Spinach Leaf Extract (SLE) attenuates LPS induced oxidative stress in liver, kidney and brain
The effects of SLE on LPS induced multiple organ injury were determined with the various oxidative stress markers in liver, kidney and brain.
LPO: The increased levels of MDA were an evident marker of lipid per oxidation. The MDA levels were increased significantly in liver (p < 0.01), kidney (p < 0.01) and brain (p < 0.001) of LPS treated mice. With SLE co-treatment significant decrease in MDA levels as compared to LPS were found as shown in (Column diagram I-III).
Catalyse activity: Catalase activity was found to be increased significantly in liver (p < 0.001), kidney (p < 0.05) and brain (p < 0.05) after LPS exposure. However co-treatment was able to reverse the effect induced by LPS significantly as shown in (Column diagram I-III).
Column diagram I
| Column Diagram I: Effect of SLE on LPS-induced changes in oxidative stress in liver on day 21: Data is mean ± S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman -Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman -Keuls test when the values of LPS + SLE are compared with LPS treated group. | ||||
| Liver | ||||
| Groups ? Biochemical? |
Control I | LPS II | LPS + SLE III | SLE IV |
| MDA(μmol/mg protein) | 0.164 ± 0.036 | 0.564 ± 0.146c | 0.385 ± 0.127ax | 0.161± 0.046y |
| Catalase(nmol of H2O2hydrolyzed/min/mg protein) | 3.83 ± 0.14 | 8.06 ± 0.88c | 5.16 ± 0.34ay | 1.34 ± 0.43ay |
| GSH(nmol GSH/mg of protein) | 0.27 ± 0.01 | 0.128 ± 0.01b | 0.21 ± 0.033x | 0.24 ±3.45 |
| GPx (nmol of NADPH used/min/mg of protein) | 1.92 ± 0.27 | 3.58 ± 0.17b | 3.01 ± 0.92 | 2.51 ± 0.13y |
Column diagram II
| Column diagram II: Effect of SLE on LPS-induced changes in oxidative stress in kidney on day 21: Data is mean ± S.D of 6-8 animals. ap <0.05, bp <0.01, c p<0.001 by Newman -Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman -Keuls test when the values of LPS + SLE are compared with LPS treated group. | ||||
| Kidney | ||||
| Groups ? Biochemical? |
Control I | LPS II | LPS + SLE III | SLE IV |
| MDA (μmol/mg protein) | 2.07 ± 1.27 | 6.33 ± 1.61b | 4.98 ± 0.78y | 1.85 ± 0.59z |
| Catalase (nmol of H2O2 hydrolyzed/min/mg protein) | 2.83 ± 0.144 | 6.615 ± 1.37a | 5.02 ± 1.62x | 2.12 ± 0.54y |
| GSH (nmol GSH/mg of protein) | 0.25 ± 0.06 | 0.13 ± 0.05b | 0.127 ± 0.01a | 0.24 ± 0.02ay |
| GPx (nmol of NADPH used/min/mg of protein) | 2.96 ± 0.25 | 5.02 ± 0.16b | 2.32 ± 0.78y | 2.94 ± 0.22y |
Column diagram III
| Column diagram III: Effect of SLE on LPS-induced changes in oxidative stress in brain on day 21: Data is mean ± S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman -Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman -Keuls test when the values of LPS + SLE are compared with LPS treated group. | ||||
| Brain | ||||
| Groups ? Biochemical? |
Control I | LPS II | LPS + SLE III | SLE IV |
| MDA (μmol/mg protein) | 0.304 ± 0.1 | 0.985 ± 0.114c | 0.612 ± 0.192ax | 0.304 ± 0.06z |
| Catalase (nmol of H2O2hydrolyzed/ min/mg protein) |
0.25 ± 0.07 | 1.41 ± 0.51b | 0.563 ± 0.150ax | 0.69 ± 0.10bx |
| GSH (nmol GSH/mg of protein) | 0.19 ± 0.01 | 0.08 ± 0.03b | 0.141 ± 0.0165bz | 0.34 ± 0.03by |
| GPx (nmol of NADPH used/min/mg of protein) | 1.07± 0.12 | 3.15 ± 0.27c | 2.148 ± 0.79cy | 1.86 ± 0.18bz |
Column diagram Column diagram IV
| Column diagram IV:Effect of SLE treatment on cognitive functions of mice subjected to LPS as measured by active avoidance test and passive avoidance test. Data is mean ± S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman-Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman-Keuls test when the values of LPS + SLE are compared with LPS treated group. | ||||
| Groups? Estimation? |
Control | LPS | LPS + SLE | SLE |
| Active avoidance (Step up latency in sec) |
3.7 ± 0.11 | 6.2 ± 0.21c | 5.3 ± 0.33y | 3.5 ± 0.25 |
| Passive avoidance (Step up latency in sec) |
65.33 ± 7.31 | 30.46 ± 6.3c | 45.49 ± 10.77z | 52.19 ± 8.6 |
GSH levels: A significant decrease (p <0.01) in GSH levels in three of the organs were observed after the mice subjected to LPS which was significantly increased following Simultaneous Treatment with SLE (Column diagram I-III).
GPX activity: Further, there was significant increase (p <0.01) in GPx activity in three of the organs after mice were subjected to LPS which was decreased significantly in SLE treated group (Column diagram I-III).
SLE attenuates LPS-induced increase in pro-inflammatory cytokines in liver
The level of liver cytokines was measured by their mRNA expression and result is shown in figure 1a. The mRNA expression of IL-1β (p <0.05) and TNF-a (p < 0.001) increased significantly in LPS treated group as compared to control group as shown in fig Ib. The expression was found to be decreased significantly with co-treatment of SLE when compared to the control group (Figure 2b).
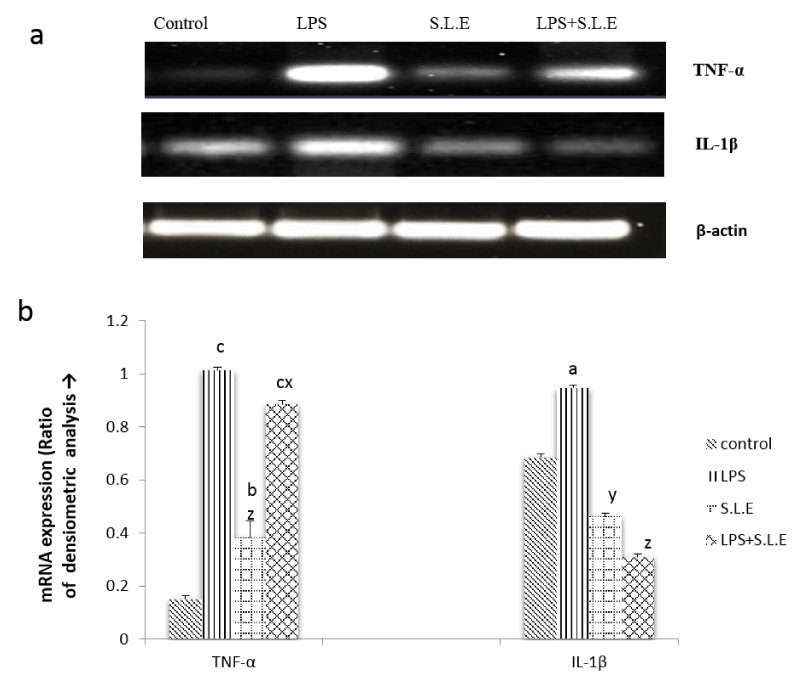 Figure 1: (a) The effect of SLE treatment on LPS induced alterations in mRNA expression of TNF-a and IL-1β. (b)Histogram showing mRNA expression of TNF-a and IL-1β in different group: Data is mean ± S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman -Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman -Keuls test when the values of LPS + SLE are compared with LPS treated group.
Figure 1: (a) The effect of SLE treatment on LPS induced alterations in mRNA expression of TNF-a and IL-1β. (b)Histogram showing mRNA expression of TNF-a and IL-1β in different group: Data is mean ± S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman -Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman -Keuls test when the values of LPS + SLE are compared with LPS treated group.
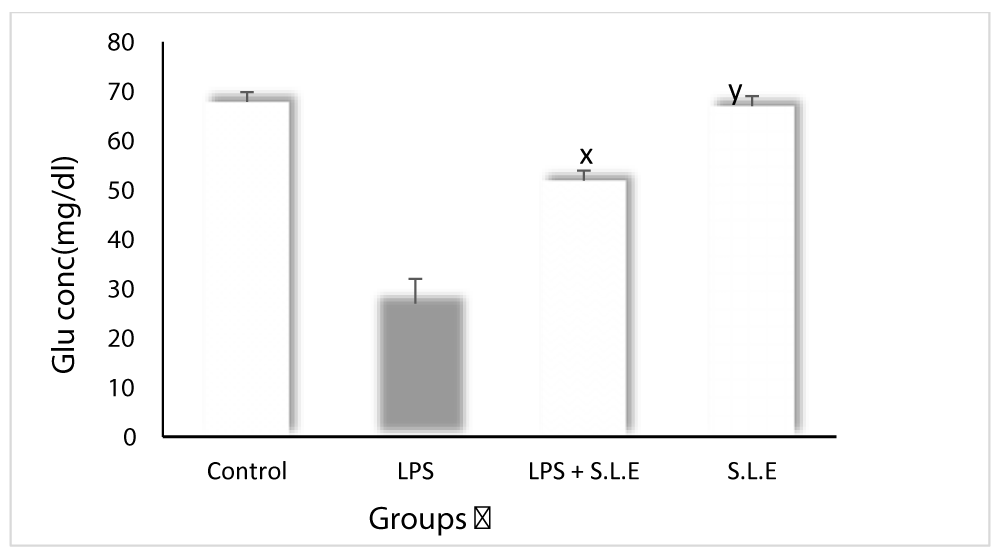 Figure 2: The effect of SLE treatment on LPS induced change in blood glucose levels: Data is mean � S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman-Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman-Keuls test when the values of LPS + SLE are compared with LPS treated group.
Figure 2: The effect of SLE treatment on LPS induced change in blood glucose levels: Data is mean � S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman-Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman-Keuls test when the values of LPS + SLE are compared with LPS treated group.
SLE normalise low blood glucose levels (hypoglycaemia) induced by LPS
Glucose levels were found to decrease significantly (p < 0.05) in LPS treated animals as compared to control but SLE normalise the level of glucose significantly as shown in (Figure 2).
SLE reverses LPS-induced histological changes in Liver, kidney and brain
LPS induced inflammation result in histological changes in liver, kidney and brain. An increase in inflammatory cells and darkly stained cells in liver (Figure 3C,D). Spinach attenuate LPS-induced histological changes as shown in figure 3-E,F). In kidney glomerular shrinkage, reduction in PCT lumen was observed in LPS treated animals as in figure 4B. SLE prevented the histological changes by LPS infection as shown in figure 4. Our histopathological analysis showed serious neuronal degeneration in LPS treated mouse brain. Darkly stained, shrunken, neurons in hippocampus were found (Figure 5C,D). Spinach inhibits the damage as shown in (Figure 5- E,F).
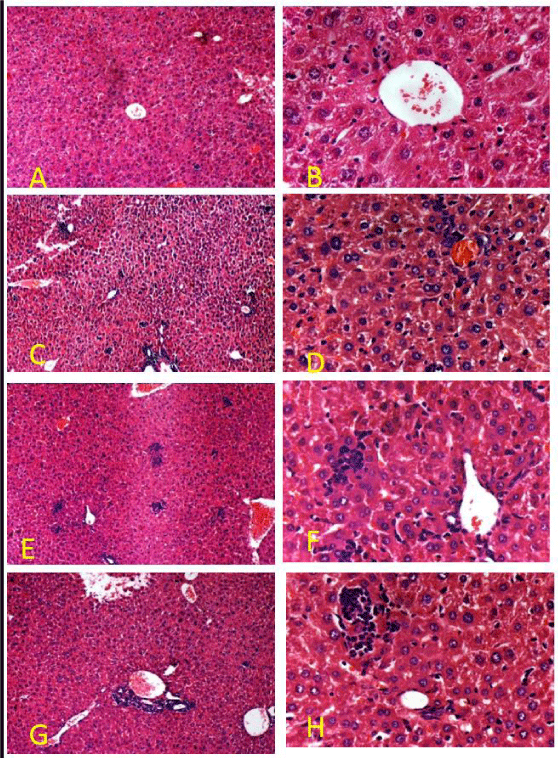 Figure 3: Representative photomicrograph of mice liver by H& E staining: (A,B) Liver of control mice at 100X and 400X respectively; (C,D) Liver of LPS treated group at 100X and 400 X respectively showed necrotic cells; (D,E) SLE co-treated along with LPS at 100 X and 400X respectively showed less necrotic cells; (F,G) SLE alone group at 100X and 400 X respectively showed normal histology.
Figure 3: Representative photomicrograph of mice liver by H& E staining: (A,B) Liver of control mice at 100X and 400X respectively; (C,D) Liver of LPS treated group at 100X and 400 X respectively showed necrotic cells; (D,E) SLE co-treated along with LPS at 100 X and 400X respectively showed less necrotic cells; (F,G) SLE alone group at 100X and 400 X respectively showed normal histology.
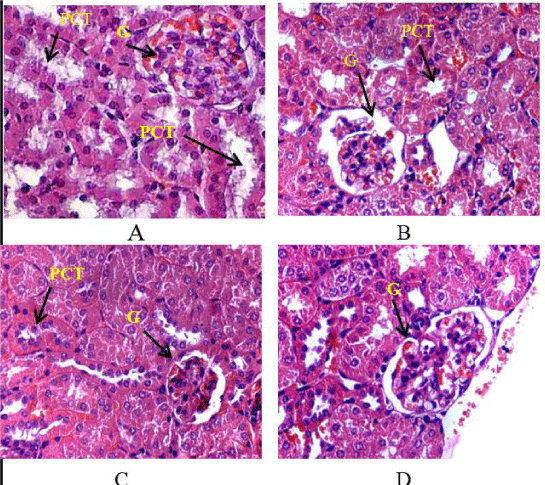 Figure 4: Representative photomicrograph of mice kidney by H& E staining: (A) Kidney of control mice at 400X; (B) kidney of LPS treated group at 400 X showed glomerulus shrinkage; (C) SLE co-treated along with LPS at 400X showed marked improvement; (D) SLE alone group at 100X and 400 X respectively showed normal histology.PCT: proximal convoluted tubule; G: Glomerulus.
Figure 4: Representative photomicrograph of mice kidney by H& E staining: (A) Kidney of control mice at 400X; (B) kidney of LPS treated group at 400 X showed glomerulus shrinkage; (C) SLE co-treated along with LPS at 400X showed marked improvement; (D) SLE alone group at 100X and 400 X respectively showed normal histology.PCT: proximal convoluted tubule; G: Glomerulus.
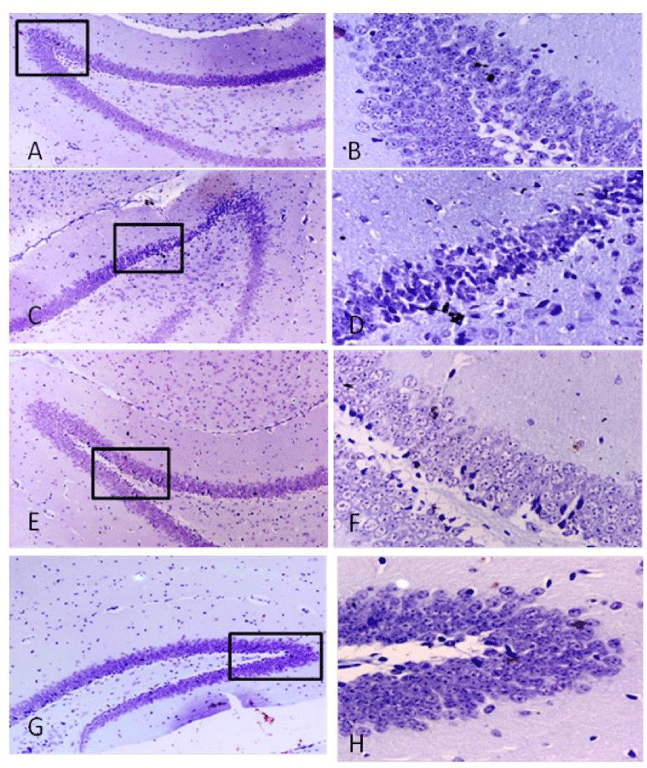 Figure 5: Representative photomicrograph of hippocampal area of mice brain by cresyl violet staining: (A,B) Brain of control mice at 100X and 400X respectively; (C,D) Brain of LPS treated group at 100X and 400 X respectively showed darkly stained and shrunken neurons; (D,E) SLE co-treated along with LPS at 100 X and 400X respectively showed very less darkly stained cells; (F,G) SLE alone group at 100X and 400 X respectively showed normal histology.
Figure 5: Representative photomicrograph of hippocampal area of mice brain by cresyl violet staining: (A,B) Brain of control mice at 100X and 400X respectively; (C,D) Brain of LPS treated group at 100X and 400 X respectively showed darkly stained and shrunken neurons; (D,E) SLE co-treated along with LPS at 100 X and 400X respectively showed very less darkly stained cells; (F,G) SLE alone group at 100X and 400 X respectively showed normal histology.
The effect of SLE on LPS induced behavioural changes
Active avoidance test used to assess learning and memory. Our results showed progressive impairment in learning and memory in LPS treated mice with respect to control. SLE co-administration with LPS proved beneficial and the animals escaped on an average of 5.3 trials as compared to LPS treated animals which exhibited 6.2 escape trials as shown in Column diagram IV. Short term memory loss was evaluated by passive avoidance test and showed a significant decrease in retention time in LPS exposed and there was a marked improvement in conjunctive treated mice (p < 0.001).
The effect of SLE on LPS induced change in acetylcholine esterase level
There was significant increase in acetylcholine esterase level after LPS exposure which related with the behavioural changes. Spinach treatment significantly (p < 0.05) reduce the level of this enzyme (Figure 6).
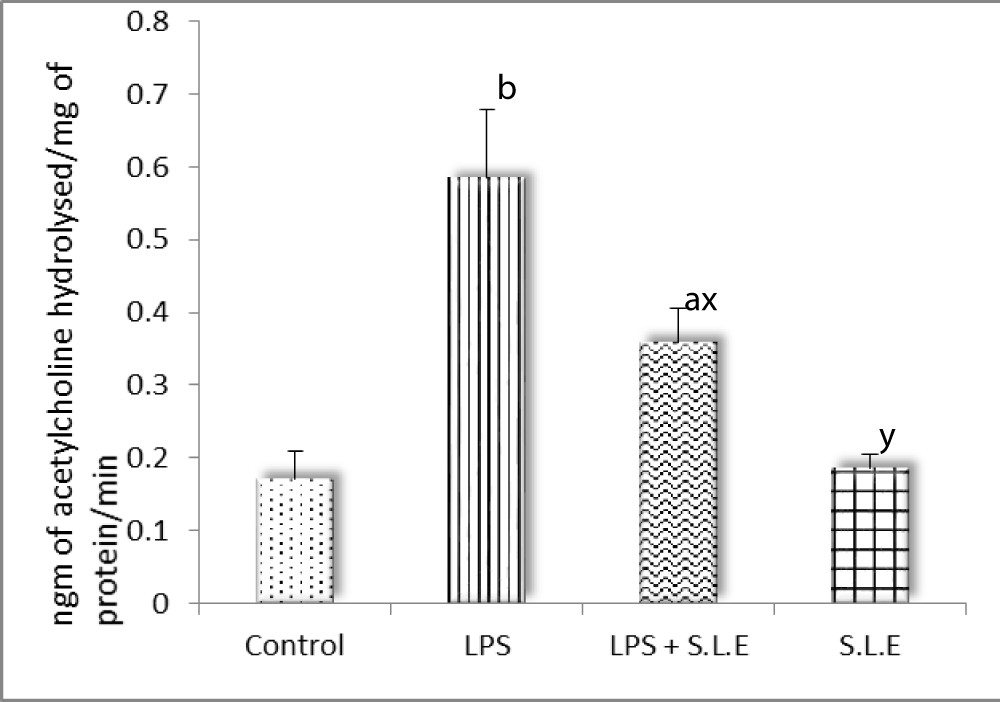 Figure 6: The effect of SLE on LPS induced change in acetylcholine esterase level: Data is mean ± S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman -Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman -Keuls test when the values of LPS + SLE are compared with LPS treated group.
Figure 6: The effect of SLE on LPS induced change in acetylcholine esterase level: Data is mean ± S.D of 6-8 animals. ap <0.05, bp <0.01, cp <0.001 by Newman -Keuls test when the values are compared with normal control group. xp <0.05, yp <0.01, zp <0.001 by Newman -Keuls test when the values of LPS + SLE are compared with LPS treated group.
Discussion
The present study is an attempt to evaluate the protective potentials of Spinach Leaf Extract (SLE) against multiple organ injury resulting from LPS exposure. LPS was administered intraperitoneally at a dose of 5 mg/kg. The dose selection was done according to Qin, et al. [28] and spinach leaf extract (50 mg/kg body wt. /day i.p) administered in conjunction with LPS.
Systemic administration of LPS resulted in its accumulation in liver first, since 70 % of the blood supply from intestine enters liver via portal vein and thus it acts as the first organ of defence [29]. There it recognizes the pathogen-associated molecular patterns (PAMP's) as danger signals, via pattern recognition receptors including families of the Toll-like receptors (TLRs) [30,33] which are present on the Kupfer cells in the liver and microglia cells in the brain [34]. This consequently leads to their activation [35] result in the production of pro-inflammatory cytokines i.e. TNF-a and IL-1β. This increased burden of several proinflammatory cytokines is responsible for multiple organ dysfunctions such as liver, lungs, brain and kidney [8]. Accordingly, we observed a significant increase in the liver cytokines levels as shown in figure 1a,b.
Further, macrophage activation i.e., kupffer cells and microglia cells leads to the activation of NADPH oxidise, a membrane bound enzyme present in their plasma membrane. This in turn accelerates ROS production which plays an important role in LPS-induced increase in oxidative burden [6,7]. These ROS formed i.e., superoxide anions can react readily with NO in the extracellular space to form more oxidants, ONOO˙ (peroxynitrite) which can readily cross the cell membranes and damage intracellular components in neighbouring cells and also result in lipid per oxidation [36]. Lipid per oxidation is the process of oxidative deterioration of polyunsaturated fatty acids in biological membranes which causes impaired structure integrity and impaired membrane functions [36]. The extent of lipid per oxidation was assessed by measuring the levels of Malondialdehyde (MDA), a by-product of lipid per oxidation. As a consequence compromised antioxidant enzyme system was observed in the present study with a significant increase in MDA levels of liver, kidney and brain after LPS treatment as compared to control (Column diagram I-III). These alterations were also associated with significant decrease in reduced glutathione (GSH) content (Column diagram I-III). GSH is an important antioxidant which directly reacts with free radicals such as ROS and RNS [38] and acts as a substrate for GPx to form oxidised glutathione (GSSG). Results showed significant increase in GPx activity in LPS treated mice could be in response to the large production of free radicals with the aid of GSH [39]. This is an indicative of greater utilisation of GPx to compact oxidative stress as compared to control. Thus, the increases in GPx activity suggested a compensatory response to increase in oxidative stress which was indicated by the elevated MDA levels [40,41]. Also, the increased catalyse activity was seen in LPS exposed animals as compared to control (Column diagram I-III). Catalyse enzyme is responsible for decomposition of hydrogen peroxide (H2O2). This increase in catalyse activity could be due to excessive production of H2O2 and other ROS [42,43] after LPS exposure in mice. This oxidative damage to lipid membrane indicated by increased MDA level is inturn reflected in histological picture of liver where a large number of necrotic cells were seen in LPS treated animals as shown in figure 3. Similar observations were made by Wang (2010) [44] in liver of LPS treated mice. Correspondingly, the histopathological changes in kidney of LPS treated animals indicated glomerular shrinkage along with reduction in PCT lumen (Figure 4). Knotted and co-workers [45] reported excessive intracranial vasoconstriction leading to renal failure [46] after administration of LPS (5mg/kg).
The cytokines IL-1 and TNFa produced in liver regulate glucose metabolism and may lead to hypoglycaemic effect in response to LPS. This was determined by enhanced levels of insulin, glucagon, and corticosterone [47,48]. Hence, significant decrease in glucose level following LPS infection was observed in present study as shown in figure 2. Whereas, Spinach co-administration normalized the blood glucose level in LPS treated animals as it regulates/inhibiting the expression of these pro-inflammatory cytokines i.e., IL-1β and TNF-a as reported by [49].
Systemic LPS exposure leads to functional breakdown of the BBB resulting in infiltration of granulocyte and passage of potential agents (environmental toxins, pathogens etc) into the brain thus, leading to microglia activation [34,50,51]. High systemic infections may also have deleterious effects on neuronal survival and may enhance the brain or CNS cytokine production, change behaviour, indicating a direct LPS to CNS communication via endothelial cells of brain [52]. This subsequently leads to enhanced cytokine production and increased oxidative burden in brain as reflected by elevated level of MDA, a marker of lipid per oxidation [36,39]. The results were further confirmed by histological changes in hippocampus area as shown in fig V. The level of acetylcholine esterase was found to increase significantly in LPS treated mice. This enzyme attached to the intercellular matrix of the synaptic cleft cause rapid hydrolysis of acetylcholine to choline and acetate enabling reuptake and recycling. Hippocampus, the region of learning and memory receives a strong cholinergic input and which has involvement in short term memory that may account for loss of memory [53]. A significant decrease in the activity of acetylcholine esterase was found in the co-treatment with SLE. The acetylcholine levels and histological changes are associated with memory loss which is evaluated by active passive avoidance test. The escape trials in active avoidance in LPS treated group were significantly high as compared to control; these results were in agreement with the previous study [10]. Similarly the retention time in the passive avoidance test, which is used to assess short term memory loss, was significantly decrease in LPS treated mice as compared to control [10].
Co-treatment of spinach leaves extract in LPS treated animals significantly prevented the alteration of different parameters. SLE significantly increased GSH levels by virtue of many active components such as flavonoids, which possess anti-oxidative property [11]. The antioxidants are responsible for stabilizing and scavenging free radicals, thereby inhibiting ROS production [54]. Various natural antioxidants (NAO) are related to the polyphone or flavonoids families. NAO extracted from spinach leaf extract [11] was found to be superior to the well-known antioxidants butylated hydroxytoluene and vitamin E. The prophylactic effect of SLE showed by significant decrease in lipid per oxidation products suggest that it as an efficient therapy of endotoxemia [13,14]. Also there was subsequent decrease in GPx activity and Catalyse activity [40]. Spinach significantly decreased the cytokines levels in LPS mice by means of the anti-inflammatory action of flavonoids and prevention of leukotriene production by lipoxygenase in leukocytes [15,16]. Spinach co-treatment in mice significantly improved the histopathological changes in brain, liver and kidney after LPS toxicity [55]. The reduction of LPS toxicity by SLE was also observed in a rat model of endotoxemia [56]. The neuroprotective efficacy of spinach extract showed by significant improvement in active passive avoidance test and decreased levels of acetylcholine esterase levels. The potential to improve learning, memory and general cognitive ability [17,57] by dietary flavonoids intake is associated with better cognitive evolution, i.e. the preservation of cognitive performance with ageing [56]. Also several studies have revealed the neuroprotective efficacy of flavonoids in improving the locomotors activity [17]. Flavonoids have been observed to attenuate NO, IL-1β and TNF-a production in activated microglia cells [58]. It has been shown by Chen and co-workers [59] that ?avonol quercetin attenuate microglia mediated neuroin?ammation via mechanisms that include inhibition of: (1) cyclooxygenase (COX-2) and iNOS expression, (2) cytokine release, and (3) NADPH oxidase activation and subsequent reactive oxygen species generation, in atrocities and microglia. Flavonoids may exert these effects via direct modulation of protein and lipid kinase signalling pathways [57], for example via the inhibition of MAPK signalling cascades, such as p38 or ERK1/2 which regulate both TNF-a and iNOS expression in activated glial cells [60].
Conclusion
Present study suggests that a single injection of LPS to mice induces a variety of inflammatory and histological changes in multiple organs [8,61,62]. The beneficial effect of Spinach leaf extract, in preventing multiple organ dysfunction and histopathological alterations in the liver, kidney and brain of mice treated with LPS, could be related to its anti-inflammatory and antioxidant properties, such as scavenging of free radicals or through inhibiting production of the chemotaxic agents [63,64].
References
- Hattori M, Taylor TD. The Human Intestinal Microbiome: A New Frontier of Human Biology. DNA Res. 2009; 16: 1-12. https://goo.gl/Tjqk5a
- Robinson CJ, Bohannan BJM, Young VB. From structure to function: the ecology of host-associated microbial communities. Microbiol Mol Biol Rev. 2010; 74: 453-76. https://goo.gl/dY1S9v
- Gillevet P, Sikaroodi M, Keshavarzian A, Mutlu EA. Quantitative Assessment of the Human Gut Microbiome Using Multitag Pyrosequencing. Chem Biodivers. 2010; 7: 1065-75. https://goo.gl/fDYjtk
- Elin RJ, Wolff SM. Biology of Endotoxin. Annual Review of Medicine. 1976; 27: 127-141. https://goo.gl/DpCjR4
- Szabo G, Bala S, Petrasek J, Gattu A. Gut-Liver Axis and Sensing Microbes. Dig Dis. 2010; 28: 737-44. https://goo.gl/ah4139
- Ben-Shaul V, Sofer Y, Bergman M, Zurovsky Y, Grossman S. Lipopolysaccharide-induced oxidative stress in the liver: comparison between rat and rabbit. Shock. 1999; 12: 288-93. https://goo.gl/aRp1ns
- Thakur V, Pritchard MT, McMullen MR, Wang Q, Nagy LE. Chronic ethanol feeding increases activation of NADPH oxidase by lipopolysaccharide in rat Kupffer cells: role of increased reactive oxygen in LPS-stimulated ERK1/2 activation and TNF-production J Leukoc Biol. 2006; 79: 1348-56. https://goo.gl/9X74TS
- Jung YY, Nam Y, Park YS, et al. Protective effect of phosphatidylcholine on lipopolysaccharide-induced acute inflammation in multiple organ injury. Korean J Physiol Pharmacol. 2013; 17: 209-16. https://goo.gl/LZNnyL
- Cunningham PN, Dyanov HM, Park P, Wang J, Newell KA, Quigg RJ. Acute renal failure in endotoxemia is caused by TNF acting directly on TNF receptor-1 in kidney. J Immunol. 2002; 168: 5817-23. https://goo.gl/L2egkJ
- Lee JW, Lee YK, Yuk DY, Choi DY, Ban SB, Oh KW, et al. Neuro-inflammation induced by lipopolysaccharide causes cognitive impairment through enhancement of beta-amyloid generation. J Neuroinflammation. 2008; 5: 37. https://goo.gl/fr1LBH
- Lomnitski L, Bergman M, Nyska A, Ben-Shaul V, Grossman S. Composition, Efficacy, and Safety of Spinach Extracts. Nutr Cancer. 2003; 46: 222-31. https://goo.gl/7SL7yt
- PINSKY A, GROSSMAN S, TROP M. LIPOXYGENASE CONTENT AND ANTIOXIDANT ACTIVITY OF SOME FRUITS AND VEGETABLES. J Food Sci. 1971; 36: 571-572. https://goo.gl/ipX5nG
- Salah N, Miller NJ, Paganga G, Tijburg L, Bolwell GP, Rice-Evans C. Polyphenolic flavanols as scavengers of aqueous phase radicals and as chain-breaking antioxidants. Arch Biochem Biophys. 1995; 322: 339-46. https://goo.gl/prz342
- van Acker SA, Tromp MN, Haenen GR, van der Vijgh WJ, Bast A. Flavonoids as scavengers of nitric oxide radical. Biochem Biophys Res Commun. 1995; 214: 755-9. https://goo.gl/fTTpG9
- Hope WC, Welton AF, Fiedler-Nagy C, Batula-Bernardo C, Coffey JW. In vitro inhibition of the biosynthesis of slow reacting substance of anaphylaxis (SRS-A) and lipoxygenase activity by quercetin. Biochem Pharmacol. 1983; 32: 367-71. https://goo.gl/9fwBej
- Kimura Y, Okuda H, Arichi S. Effects of baicalein on leukotriene biosynthesis and degranulation in human polymorphonuclear leukocytes. Biochim Biophys Acta. 1987; 922: 278-86. https://goo.gl/eDdqXw
- Vauzour D, Vafeiadou K, Rodriguez-Mateos A, Rendeiro C, Spencer JP. The neuroprotective potential of flavonoids: a multiplicity of effects. Genes Nutr. 2008; 3: 115-26. https://goo.gl/FPyp8T
- Bergman M, Varshavsky L, Gottlieb HE, Grossman S. The antioxidant activity of aqueous spinach extract: chemical identification of active fractions. Phytochemistry. 2001; 58: 143-52. https://goo.gl/SW7RxY
- Miyamoto M, Shintani M, Nagaoka A, Nagawa Y. Lesioning of the rat basal forebrain leads to memory impairments in passive and active avoidance tasks. Brain Res. 1985; 328: 97-104. https://goo.gl/XEqKdu
- LOWRY OH, ROSEBROUGH NJ, FARR AL, RANDALL RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951; 193: 265-75. https://goo.gl/35fE3j
- Wills ED. Mechanisms of lipid peroxide formation in animal tissues. Biochem J. 1966; 99: 667-76. https://goo.gl/PrcWVK
- Luck. Catalase, in Methods of Enzymatic Analysis. In: H. W. Bergmeyer E (ed) Acad. Press. New York, NY, USA. 1963; 3: 885-894. https://goo.gl/YQN8Ff
- ELLMAN GL. Tissue sulfhydryl groups. Arch Biochem Biophys. 1959; 82: 70-7. https://goo.gl/622pQc
- Paglia DE, Valentine WN. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med. 1967; 70: 158-69. https://goo.gl/bD19uz
- G E Histochemistry PA, Pearse AG. This Week's Citation Classic. R Postgrad Mcd. Sch . 1960; 998.
- Presnell JK, Schreibman MP, Humason GL. Humason's Animal tissue techniques. Johns Hopkins University Press. 1997: 572. https://goo.gl/pqX4Mb
- KELLER GJ. A reliable Nissl method. Bull Int Assoc Med Mus. 1945; 25: 77. https://goo.gl/YfcJ8r
- Qin L, Wu X, Block ML, Liu Y, Breese GR, Hong JS, et al. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia. 2007; 55: 453-62. https://goo.gl/YKbG5y
- Son G, Kremer M, Hines IN. Contribution of gut bacteria to liver pathobiology. Gastroenterology Research and Practice. 2010: 13. https://goo.gl/GLNWbx
- Miller MA, McTernan PG, Harte AL, Silva NF, Strazzullo P, Alberti KG, et al. Ethnic and sex differences in circulating endotoxin levels: A novel marker of atherosclerotic and cardiovascular risk in a British multi-ethnic population. Atherosclerosis. 2009; 203: 494-502. https://goo.gl/hmRGX2
- Sun L, Yu Z, Ye X, Zou S, Li H, Yu D et al. A Marker of Endotoxemia Is Associated With Obesity and Related Metabolic Disorders in Apparently Healthy Chinese. Diabetes Care. 2010; 33: 1925-32. https://goo.gl/pc7Twh
- Benson S, Kattoor J, Wegner A, Hammes F, Reidick D, Grigoleit JS, et al. Acute experimental endotoxemia induces visceral hypersensitivity and altered pain evaluation in healthy humans. Pain. 2012; 153: 794-9. https://goo.gl/ZjZH2o
- Moreira AP, Texeira TF, Ferreira AB, Peluzio Mdo C, Alfenas Rde C. Influence of a high-fat diet on gut microbiota, intestinal permeability and metabolic endotoxaemia Br J Nutr. 2012; 108: 801-9. https://goo.gl/3MQJRf
- Kloss CU, Bohatschek M, Kreutzberg GW, Raivich G. Effect of Lipopolysaccharide on the Morphology and Integrin Immunoreactivity of Ramified Microglia in the Mouse Brain and in Cell Culture. Exp Neurol. 2001; 168: 32-46. https://goo.gl/Ya82fC
- Beutler B, Rietschel ET. imeline: Innate immune sensing and its roots: the story of endotoxin. Nat Rev Immunol. 2003; 3: 169-76. https://goo.gl/ngju2C
- Ferihan Cetin F, Dincer S, Ay R, Guney S. Systemic taurine prevents brain from lipopolysaccharide-induced lipid peroxidation in rats. African J Pharm Pharmacol. 2012; 6: 1099-1105. https://goo.gl/efDt65
- Gutteridge JMC. Lipid Peroxidation and Antioxidants as Biomarkers of Tissue Damage. Clin Chem. 1995; 41: 1819-28. https://goo.gl/JjGSnX
- Luperchio S, Tamir S, Tannenbaum SR. NO-induced oxidative stress and glutathione metabolism in rodent and human cells. Free Radic Biol Med. 1996; 21: 513-9. https://goo.gl/wpNQbe
- Abd el-gawad HM, Khalifa AE. Quercetin, Coenzyme Q10, and l -canavanine as protective agents against lipid peroxidation and nitric oxide generation in endotoxin-induced shock in rat brain. Pharmacol Res. 2001; 43: 257-63. https://goo.gl/4gZ4qe
- Ben-Shaul V, Lomnitski L, Nyska A, Carbonatto M, Peano S, Zurovsky Y, et al. Effect of natural antioxidants and apocynin on LPS-induced endotoxemia in rabbit. Hum Exp Toxicol. 2000; 19: 604-14. https://goo.gl/oJTmbc
- ZHU Y, CARVEY P, LING Z. Altered glutathione homeostasis in animals prenatally exposed to lipopolysaccharide. Neurochem Int. 2007; 50: 671-80. https://goo.gl/BqzS3L
- Zmijewski JW, Lorne E, Zhao X, Tsuruta Y, Sha Y, Liu G, et al. Antiinflammatory Effects of Hydrogen Peroxide in Neutrophil Activation and Acute Lung Injury. Am J Respir Crit Care Med. 2009; 179: 694-704. https://goo.gl/ZsFPXd
- Zoccarato F, Toscano P, Alexandre A. Dopamine-derived Dopaminochrome Promotes H2O2 Release at Mitochondrial Complex I. J Biol Chem. 2005; 280: 15587-94. https://goo.gl/BhwHqp
- Wang K, Damjanov I, Wan Y-JY. The protective role of pregnane X receptor in lipopolysaccharide/D-galactosamine-induced acute liver injury. Lab Invest. 2010; 90: 257-65. https://goo.gl/86Fy82
- Knotek M, Rogachev B, Wang W, Ecder T, Melnikov V, Gengaro PE, et al. Endotoxemic renal failure in mice: Role of tumor necrosis factor independent of inducible nitric oxide synthase. Kidney Int. 2001; 59: 2243-9. https://goo.gl/jzA8Tc
- Epstein FH, Parrillo JE. Pathogenetic Mechanisms of Septic Shock. N Engl J Med. 1993; 328: 1471-7. https://goo.gl/Va2Nr1
- Vogel SN, Henricson BE, Neta R. Roles of interleukin-1 and tumor necrosis factor in lipopolysaccharide-induced hypoglycemia. Infect Immun. 1991; 59: 2494-8. https://goo.gl/EVDEMt
- Oguri S, Motegi K, Iwakura Y, Endo Y. Primary role of interleukin-1 alpha and interleukin-1 beta in lipopolysaccharide-induced hypoglycemia in mice. Clin Diagn Lab Immunol. 2002; 9: 1307-12. https://goo.gl/tNNb8a
- Santangelo C, Vari R, Scazzocchio B, Di Benedetto R, Filesi C, Masella R. Polyphenols, intracellular signalling and inflammation. Ann Ist Super Sanita. 2007; 43: 394-405. https://goo.gl/L7hr3h
- Bohatschek M, Werner A, Raivich G. Systemic LPS Injection Leads to Granulocyte Influx into Normal and Injured Brain: Effects of ICAM-1 Deficiency. Exp Neurol. 2001; 172: 137-52. https://goo.gl/K2tZzt
- Cardoso FL, Kittel A, Veszelka S, Palmela I, Toth A, Brites D, et al. Exposure to Lipopolysaccharide and/or Unconjugated Bilirubin Impair the Integrity and Function of Brain Microvascular Endothelial Cells. PLoS One. 2012; 7: e35919. https://goo.gl/kiXtHd
- Murray CL, Skelly DT, Cunningham C. Exacerbation of CNS inflammation and neurodegeneration by systemic LPS treatment is independent of circulating IL-1β and IL-6. J Neuroinflammation. 2011; 8: 50. https://goo.gl/1Kd5vK
- Das A, Shanker G, Nath C, Pal R, Singh S, Singh H. A comparative study in rodents of standardized extracts of Bacopa monniera and Ginkgo biloba: anticholinesterase and cognitive enhancing activities. Pharmacol Biochem Behav. 2002; 73: 893-900. https://goo.gl/MVjXDU
- Niki E. Action of Antioxidants against Oxidative Stress. Advances in DNA Damage and Repair. Springer US, Boston, MA, 1999: 313-318. https://goo.gl/SYsYzG
- Lomnitski L, Foley JE, Grossman S, Shaul VB, Maronpot RR, Moomaw CR, et al. Effects of Apocynin and Natural Antioxidant from Spinach on Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Induction in Lipopolysaccharide-Induced Hepatic Injury in Rat. Pharmacol Toxicol. 2000; 87: 18-25. https://goo.gl/XVa8Tc
- Letenneur L, Proust-Lima C, Le Gouge A, Dartigues JF, Barberger-Gateau P. Flavonoid Intake and Cognitive Decline over a 10-Year Period. Am J Epidemiol. 2007; 165: 1364-71. https://goo.gl/2rQNtz
- Spencer JP. Flavonoids: modulators of brain function?. Br J Nutr. 2008; 99: 60-77. https://goo.gl/9WJ7CP
- Lau FC, Shukitt-Hale B, Joseph JA. Nutritional intervention in brain aging: reducing the effects of inflammation and oxidative stress. Subcell Biochem. 2007; 42: 299-318. https://goo.gl/aetrh1
- Chen JJ, Swope DM. Clinical Pharmacology of Rasagiline: A Novel, Second-Generation Propargylamine for the Treatment of Parkinson Disease. J Clin Pharmacol. 2005; 45: 878-894. https://goo.gl/Jo9jnQ
- Bhat NR, Zhang P, Lee JC, Hogan EL. Extracellular signal-regulated kinase and p38 subgroups of mitogen-activated protein kinases regulate inducible nitric oxide synthase and tumor necrosis factor-alpha gene expression in endotoxin-stimulated primary glial cultures. J Neurosci. 1998; 18:1633-41. https://goo.gl/x3vpcW
- Van Deventer SJ, ten Cate JW, Tytgat GN. Intestinal endotoxemia. Clinical significance. Gastroenterology. 1988; 94: 825-31. https://goo.gl/ByCpu8
- Wenzel RP, Pinsky MR, Ulevitch RJ, Young L. Current Understanding of Sepsis. Clin Infect Dis. 1996; 22: 407-413.https://goo.gl/8BtdR4
- Nagy L, Lee TH, Goetzl EJ, Pickett WC, Kay AB. Complement receptor enhancement and chemotaxis of human neutrophils and eosinophils by leukotrienes and other lipoxygenase products. Clin Exp Immunol. 1982; 47: 541-7. https://goo.gl/EEWnLL
- Munoz NM, Douglas I, Mayer D, Herrnreiter A, Zhu X, Leff AR. Eosinophil chemotaxis inhibited by 5-lipoxygenase blockade and leukotriene receptor antagonism. Am J Respir Crit Care Med. 1997; 155: 1398-1403. https://goo.gl/d4M4AZ
Authors submit all Proposals and manuscripts via Electronic Form!